Clear Sky Science · sv
Effektorers konformationella plasticitet möjliggör linspecifik sekretion via Hcp-heterohexamerer i tarmens symbionter
Mikrobiella dueller i människans tarm
Våra tarmar hyser täta samhällen av mikrober som ständigt tävlar om utrymme och näring. För att överleva använder vissa bakterier molekylära vapen som kan punktera och förgifta konkurrenter och på så sätt påverka vilka arter som frodas i tarmen. Denna studie avslöjar hur en vanlig tarmbakterie, Bacteroides fragilis, skräddarsyr ett litet injektionsverktyg så att det kan leverera specifika toxiner till närliggande bakterier med anmärkningsvärd precision.
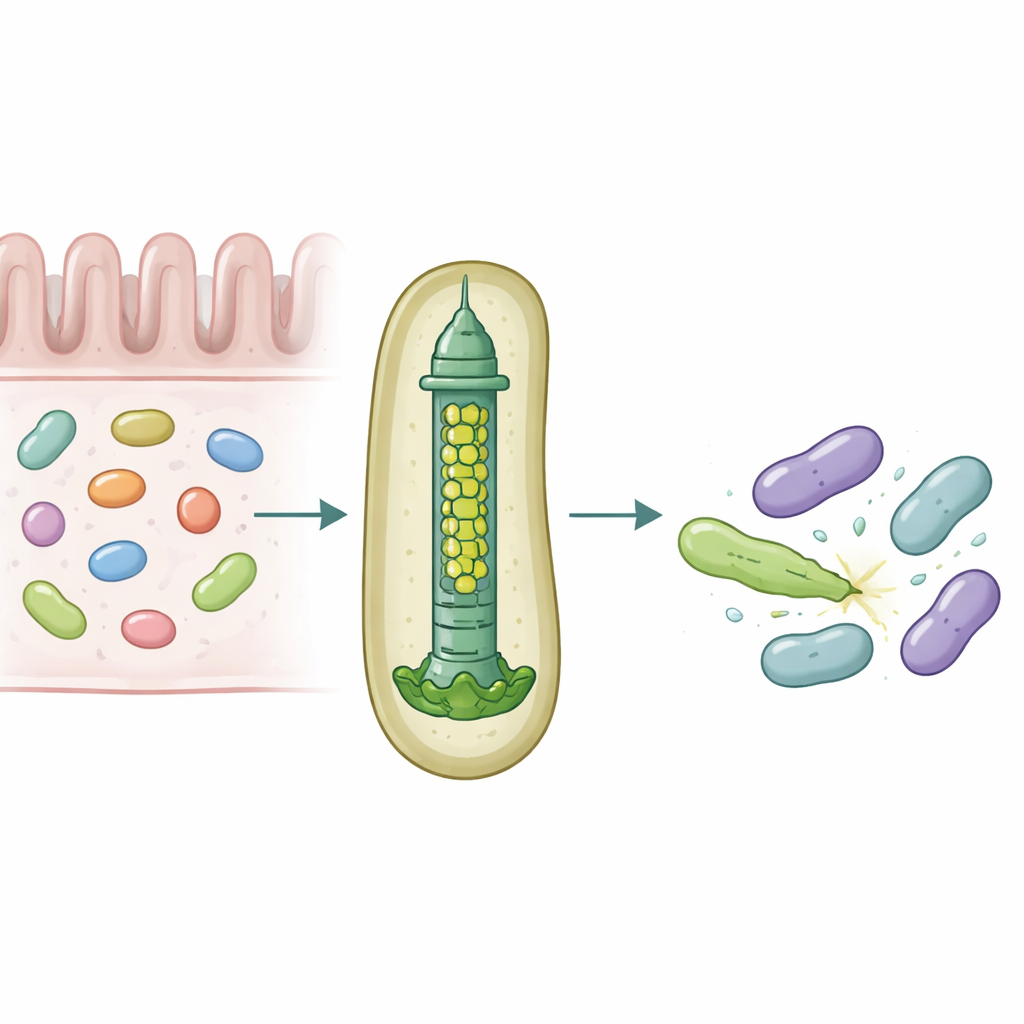
En mikroskopisk lanspistol för bakteriekrigföring
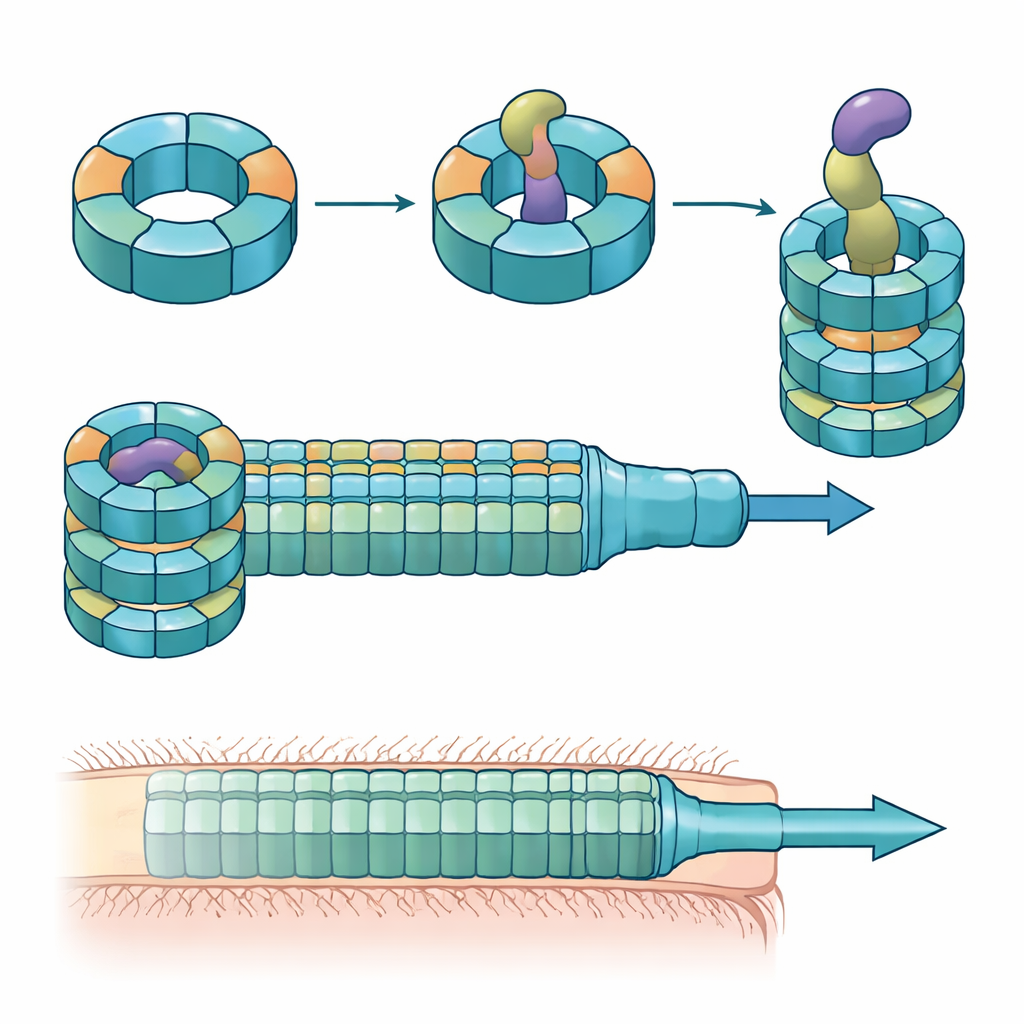
Många gramnegativa bakterier använder en anordning som kallas Typ VI sekretionssystem, en fjäderladdad struktur som fungerar som en omvänd fagtail, för att skjuta giftiga ”effektorer” in i intilliggande celler. En central del av detta system är ett rör byggt av staplade ringar av ett protein kallat Hcp. I de flesta arter finns en form av Hcp per system, men B. fragilis från människans tarm har fem olika hcp-gener i en och samma locus. Den biologiska anledningen till denna till synes överflöd var tidigare okänd, särskilt eftersom detta system är ovanligt effektivt på att döda närbesläktade bakterier i laboratorie- och djurmodeller.
Specialiserade ringar byggda av blandade delar
Forskarna kombinerade muskolonisationsförsök, blandade bakteriekulturer och proteinbiokemi för att kartlägga rollerna för dessa Hcp-variant. De fann att endast tre—Hcp1, Hcp2 och Hcp3—krävs för effektiv nedbrytning av en viss effektor kallad Bte1. Hcp1 bildar huvudröret i sekretionsmaskinen, som väntat. Hcp2 och Hcp3 gör däremot något annat: de monterar ihop sig till en blandad sexdelad ring, en heterohexamer, som inte kan bildas av något av proteinerna ensam. Med kryo-elektronmikroskopi visade teamet att dessa heterohexamerer består av fyra Hcp2- och två Hcp3-enheter ordnade i en symmetrisk ring, vilket skapar en styv por som kan fånga upp Bte1 men inte blandas med Hcp1-ringarna.
Formskiftande last och ett lås-och-nyckel-passar
Högupplösta strukturer av Hcp2–Hcp3-ringen bunden till Bte1 visade att effektorn måste omorganisera delar av sin struktur dramatiskt för att passa tätt inne i den centrala kanalen. Flera av dess α-helixar svänger för tiotals grader för att undvika kollision med proteintuben, vilket demonstrerar vad författarna kallar konformationell plasticitet. När de konstruerade en disulfidbindning för att låsa en av dessa helixar på plats kunde Bte1 inte längre binda ringen effektivt, och dess dödande aktivitet minskade i konkurrensförsök. Detaljerad kartläggning av kontaktytorna pekade ut specifika aminosyror på både Hcp2 och Hcp3 som är nödvändiga för bindning och sekretion. Att mutera bara en rest på någon av parterna räckte för att störa leveransen utan att själva ringen bröts sönder.
Medutvecklade ringar och toxiner över tarmens linjer
Teamet undersökte sedan om denna mekanism är unik för en stam eller delad över B. fragilis-linjer. Genom att jämföra Hcp2–Hcp3-par och deras närliggande effektorgener från dussintals stammar fann de att varje linje bär en karakteristisk effektor i en variabel region av locus och ett matchande Hcp2–Hcp3-par. Korsbytning av dessa par mellan stammar visade att varje blandad ring endast känner igen sin egen effektor, även om olika ringar fortfarande kan monteras strukturellt. Hcp3 innehåller i synnerhet mycket variabla ytor på ringens insida som fungerar som huvudsaklig bestämningsfaktor för vilken effektor som kan binda. Trots denna sekvensdiversitet delar effektorerna själva en bevarad N-terminal veckning som dockar in i ringen som en återanvändbar modul, medan deras C-terminala toxiska domäner varierar stort.
Varför detta formmatchningssystem är viktigt
Tillsammans skisserar dessa fynd en ”lås-och-nyckel” ko-evolutionär modell: ringens insida i Hcp2–Hcp3 fungerar som ett linjespecifikt lås, och varje effektors bevarade N-terminala modul fungerar som en nyckel som subtilt omformats i olika stammar. Effektorproteiner kan böja och omveckla sig precis så mycket att de trådar igenom ringen utan att kärva fast i röret, vilket möjliggör leverans av relativt stora eller komplexa toxiner genom en smal kanal. Detta arbete förklarar hur tarmens symbionter finjusterar sina molekylära lansar för att använda olika toxiner mot mikrobiella rivaler, och antyder att genom att omkonfigurera dessa lås-och-nyckel-gränssnitt kan forskare en dag konstruera Typ VI-system som programmerbara leveransplattformar för nyttiga molekyler i mikrobiala samhällen.
Citering: Zheng, S., Li, W., Fan, L. et al. Effector conformational plasticity enables lineage-specific secretion via Hcp heterohexamers in gut symbionts. Nat Commun 17, 2994 (2026). https://doi.org/10.1038/s41467-026-69309-z
Nyckelord: typ VI sekretionssystem, Bacteroides fragilis, bakteriell konkurrens, proteinkonformationell plasticitet, mikrobiomteknik