Clear Sky Science · ru
Квантитативная живая визуализация показывает, что PRICKLE1 контролирует морфогенез переходной зоны нервной трубки независимо от планарной клеточной полярности
Как раннее формирование нервной системы формирует наш мозг и спинной мозг
Ещё до того, как человеческий эмбрион достигает размера рисового зернышка, плоский лист клеток должен сложиться и запечататься в полую трубку, которая станет мозгом и спинным мозгом. Когда этот процесс проходит неправильно, у новорождённых могут возникать серьёзные дефекты позвоночника. В этом исследовании с использованием передовой живой визуализации в крошечных эмбрионах перепела авторы наблюдают эту трансформацию в реальном времени, раскрывая неожиданное поведение клеток и ключевой регуляторный белок, которые вместе помогают предотвращать определённый класс спинальных нарушений.
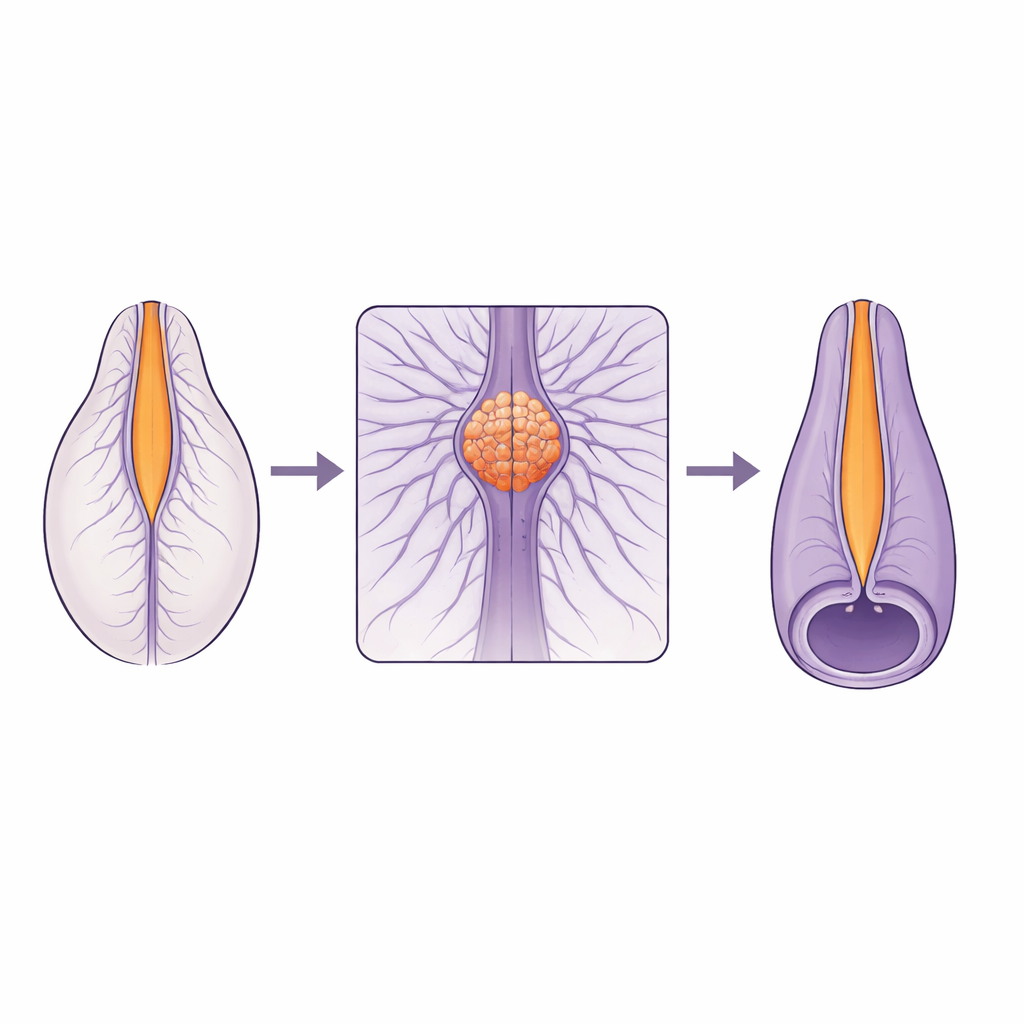
Тонкое стыковочное место в растущем позвоночнике
Спинной мозг не формируется одинаково по всей своей длине. Верхняя часть возникает путём сгибания и закрытия плоской ткани, в то время как нижняя часть формируется из сплошного столбика клеток, который образует полость изнутри. Эти две «строительные программы» должны встретиться и слиться плавно в небольшой области, называемой переходной зоной. Многие человеческие спинальные дефекты сосредоточены вблизи этой зоны, что указывает на её повышенную уязвимость. До сих пор учёным не хватало живых, высокоразрешающих изображений, позволяющих понять, как именно клетки двигаются и меняют форму в этой зоне во время закрытия трубки.
Наблюдение за движением клеток и их погружением под поверхность
Исследователи использовали трансгенных эмбрионов перепела, чьи клетки светятся под микроскопом, что позволило проследить сотни отдельных клеток в течение нескольких часов. Они обнаружили, что формирование переходной нервной трубки происходит за счёт двух скоординированных движений. Сначала клетки с боков ползут к срединной линии — движение, называемое медиолатеральной конвергенцией, которое сужает ткань. Затем полоса клеток прямо по центру совершает более драматичный шаг: эти клетки уменьшают свои верхние (апикальные) поверхности, а затем опускаются под поверхность в более глубокие слои ткани — поведение, известное как ингрессия. Эта ингрессия напоминает эпителиально‑мезенхимальный переход (ЭМТ), при котором упорядоченные пластинчатые клетки становятся более подвижными, но здесь это происходит в клетках, которые сохраняют нейральную идентичность и в основном в дальнейшем вносят вклад в спинной мозг, а не в другие ткани.
Неожиданная роль известного белка полярности
Ранее в этой переходной области был выявлен белок PRICKLE1. Обычно PRICKLE1 известен как часть системы планарной клеточной полярности (PCP), которая ориентирует клетки бок‑о‑бок и помогает удлинять ткани. Ожидалось бы, что удаление PRICKLE1 исказит эту полярность. Вместо этого, когда команда снизила уровни PRICKLE1 выборочно в переходной зоне, обычные шаблоны полярности и длинные актиновые пучки, направляющие конвергенцию, в основном сохранились. Изменилось поведение медиальных клеток: они не сократили свои апикальные поверхности, не совершили нормальной ингрессии, и заднее отверстие нервной трубки осталось незакрытым, что вызвало переходные дефекты нервной трубки в эмбрионах.
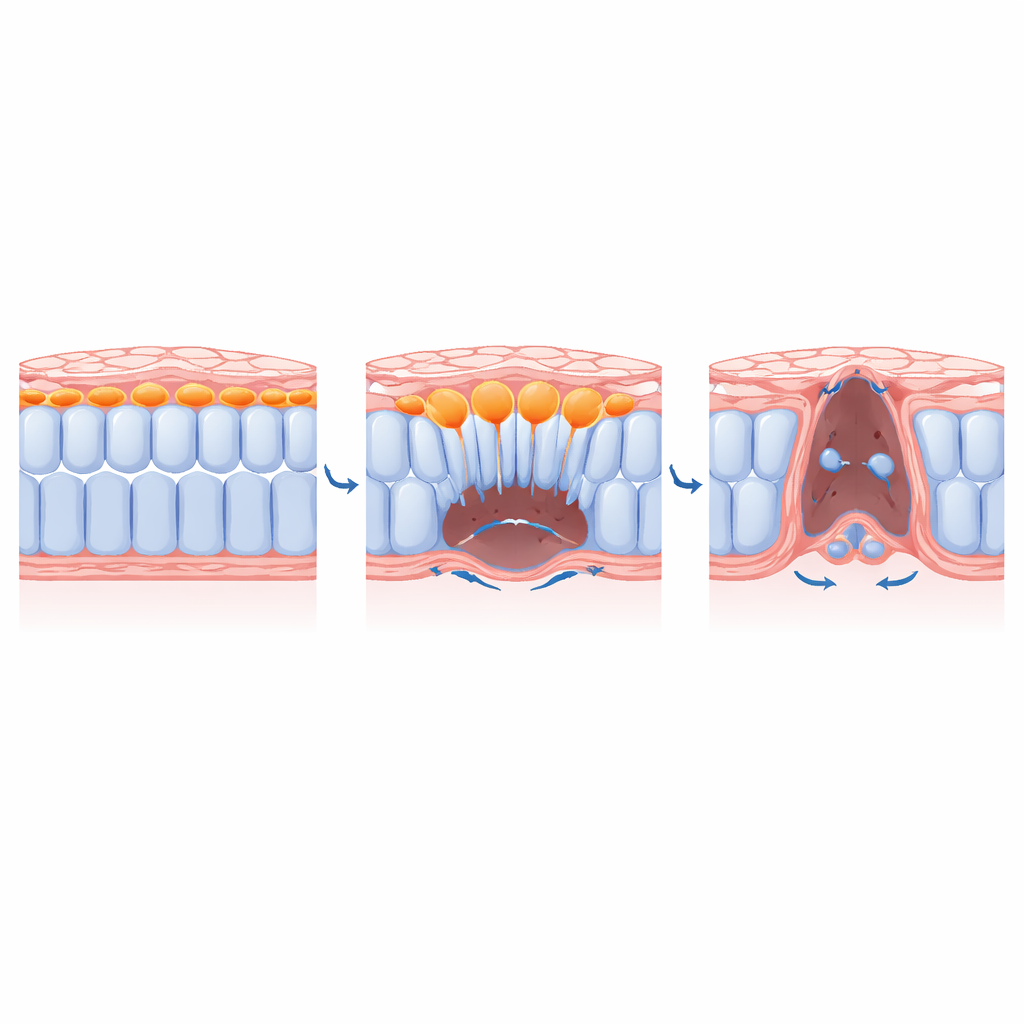
Как центральные клетки освобождают место для соседей
Комбинируя генетические инструменты, препараты и даже светозависимые ингибиторы миозина (молекулы‑мотора, обеспечивающей сокращение), команда разъяснила, как контролируется ингрессия. Они показали, что ключевые регуляторы ЭМТ, такие как транскрипционный фактор SLUG, и сигналы от факторов роста фибробластов необходимы, чтобы медиальные клетки приняли выдвижную, миграционную форму и разрушили поддерживающий матрикс под собой. Блокирование этих сигналов удерживало медиальные клетки у дорсальной поверхности и предотвращало их продвижение вентрально. Критически важно, что при отключении сокращения только в медиальных клетках эти клетки больше не ингрессировали, и боковые соседи не могли полностью сойтись к срединной линии. Другими словами, акт погружения центральных клеток физически освобождает пространство, позволяя оставшейся пластине «молнией» смыкаться.
Почему это важно для человеческих спинальных дефектов
Работа показывает, что у PRICKLE1 есть роль, независимая от PCP, на апикальной коре медиальных клеток: он способствует накоплению контрактильного актина и миозина, необходимых для апикального сжатия и ингрессии. Без этой функции, несмотря на сохранение бок‑о‑бок сигналов полярности, центральный процесс, похожий на ЭМТ, останавливается, и нервная трубка не закрывается в переходной зоне. Поскольку у людей мутации в PRICKLE1 чаще всего связаны с локализованными спинальными диспрафиями — переходными дефектами нервной трубки — эти результаты дают конкретное клеточное объяснение. Для неспециалиста основной вывод таков: небольшая группа клеток должна одновременно стянуть свои верхушки и нырнуть под поверхность в нужном месте и в нужное время, а белок PRICKLE1 является критическим переключателем для этого движения. Когда этот переключатель даёт сбой, соединение верхней и нижней частей спинного мозга не формируется правильно, что помогает объяснить специфический и серьёзный класс врождённых дефектов.
Цитирование: Wang, J.X., Alvarez, Y.D., Tan, S.Z. et al. Quantitative live imaging reveals PRICKLE1 controls junctional neural tube morphogenesis independent of Planar Cell Polarity. Nat Commun 17, 3654 (2026). https://doi.org/10.1038/s41467-026-71242-0
Ключевые слова: пороки закрытия нервной трубки, PRICKLE1, эпителиально‑мезенхимальный переход, эмбриональное развитие, миграция клеток