Clear Sky Science · pl
Przeniesienie protonu za pośrednictwem ruchomej klapki ujawnione w dekarboksylazie Fe/αKG TraH
Dlaczego mała ruchoma pętelka ma znaczenie
W komórkach mikroorganizmów wyspecjalizowane białka cicho konstruują i przekształcają złożone cząsteczki, w tym potencjalne antybiotyki. W tym badaniu autorzy skupili się na jednym z takich białek, zwanym TraH, pochodzącym z powszechnego grzyba psującego żywność. Naukowcy pokazują, że mała elastyczna „klapka” w TraH działa jak ruchoma bramka kontrolująca sposób, w jaki enzym usuwa fragment cząsteczki oraz — w niektórych przypadkach — decyduje o rodzaju zachodzącej przemiany chemicznej. Zrozumienie tej ruchomej części nie tylko pogłębia naszą wiedzę o działaniu naturalnych katalizatorów, lecz może też pomóc naukowcom w projektowaniu nowych enzymów do syntezy leków i innych cennych związków.
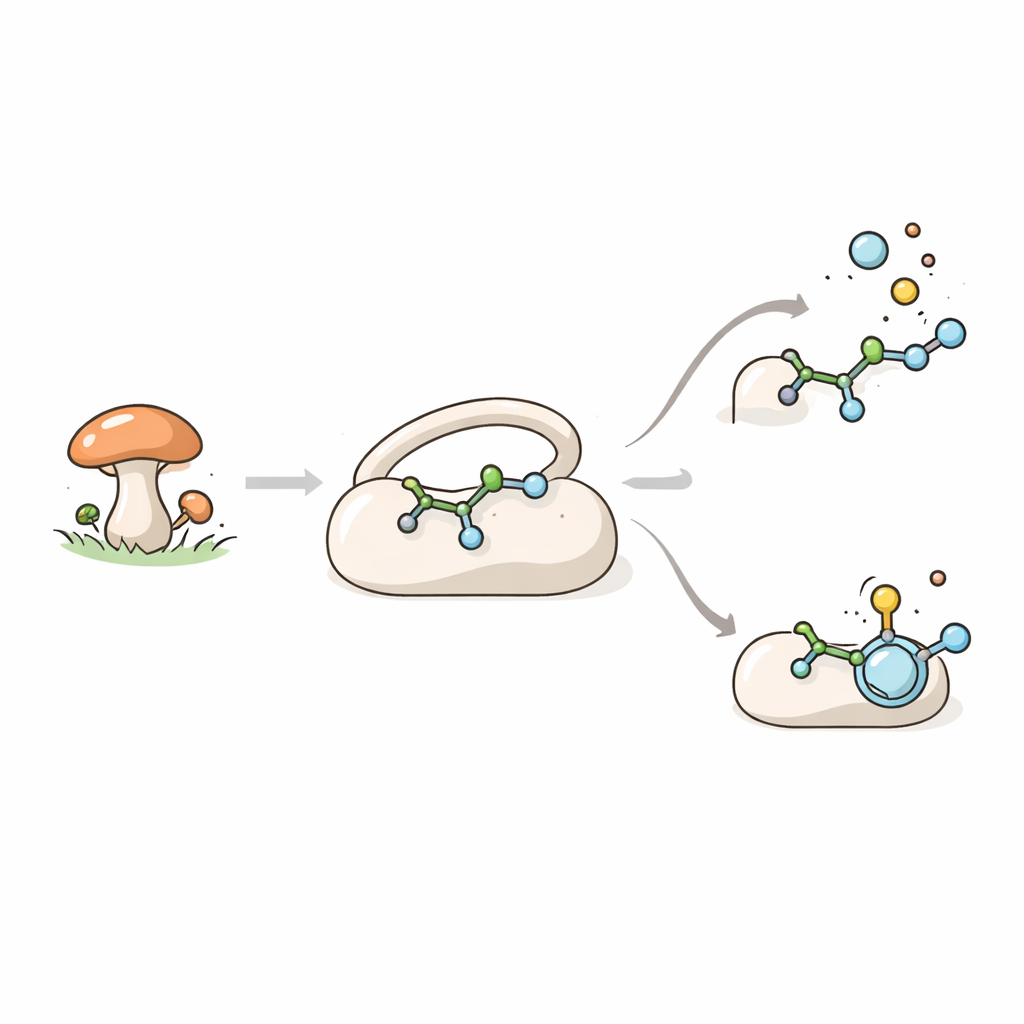
Grzybowy pomocnik w tworzeniu cząsteczek
TraH należy do dużej rodziny enzymów wykorzystujących żelazo i pomocniczą cząsteczkę α‑ketoglutaran do przeprowadzania silnych reakcji utleniania. Enzymy te odgrywają kluczową rolę w chemii mikroorganizmów, gdzie uczestniczą w powstawaniu metabolitów wtórnych, takich jak barwniki, toksyny i związki o działaniu podobnym do leków. U grzyba Penicillium crustosum TraH bierze udział w biosyntezie kwasu terrestrynowego, związku o obiecujących właściwościach przeciwbakteryjnych i przeciwwirusowych. Podstawową funkcją TraH jest odcinanie grupy karboksylowej z prekursora nazwanego kwasem crustosycznym, przy jednoczesnym wprowadzeniu podwójnego wiązania do cząsteczki. Co ciekawe, TraH może też działać na blisko spokrewnioną formę tego związku — metylowy ester — ale w tym wypadku jedynie wprowadza podwójne wiązanie, nie usuwając CO2.
Zamykająca się klapka, która przekształca miejsce aktywne
Za pomocą krystalografii rentgenowskiej autorzy uzyskali zdjęcia TraH w wysokiej rozdzielczości w różnych stanach: pustym, związanym jedynie z metalem oraz związanym z metalem i albo kwasem crustosycznym, albo jego metylowym esterem. Wszystkie formy mają wspólne sztywne jądro w kształcie podwójnej warstwowej beta‑helisy, które utrzymuje atom żelaza, gdzie zachodzi reakcja. Otaczające to jądro są dodatkowe fragmenty na końcach białka, w tym pętelka przy froncie, która zachowuje się jak klapka. W stanie niezwiązanym klapka jest wiotka i częściowo niewidoczna w strukturze krystalicznej, co wskazuje na jej wędrowanie. Gdy wiąże się prawdziwy substrat karboksylowy, klapka opada, staje się dobrze uporządkowana i wraz z pobliskimi helisami uszczelnia miejsce aktywne, tak że cząsteczka leży głęboko w zamkniętej kieszeni. Dla substratu‑estru metylowego jednak znaczna część klapki pozostaje nieuporządkowana i cofnięta, co pokazuje, że jej zamknięcie zależy od natury związanego liganda.
Przekaźnik protonowy oparty na wodzie
Bardziej szczegółowa analiza struktury z zamkniętą klapką i związanym kwasem crustosycznym ujawniła misterną sieć wiązań wodorowych łączących klapkę z konserwowanym lizyną (K191) w jądrze enzymu oraz z grupą karboksylową substratu. Kluczowe reszty w klapce pozycjonują jedną lub dwie cząsteczki wody między K191 a reaktywnym końcem substratu. Aby sprawdzić znaczenie tej sieci, zespół wprowadził pojedyncze mutacje aminokwasowe w klapce i pobliskim jądrze. Mutacje niszczące sieć, szczególnie w pozycjach Q99, D112, K114, K191 i Y214, silnie osłabiły lub zablokowały reakcję dekarboksylacji, podczas gdy zmiany w sąsiednich resztach niebiorących udziału w sieci miały niewielki efekt. Natomiast większość tych destrukcyjnych mutacji niemal nie wpływała na zdolność enzymu do desaturacji substratu‑estru metylowego, co sugeruje, że sieć pośredniczona przez wodę jest kluczowa specyficznie dla utraty dwutlenku węgla.
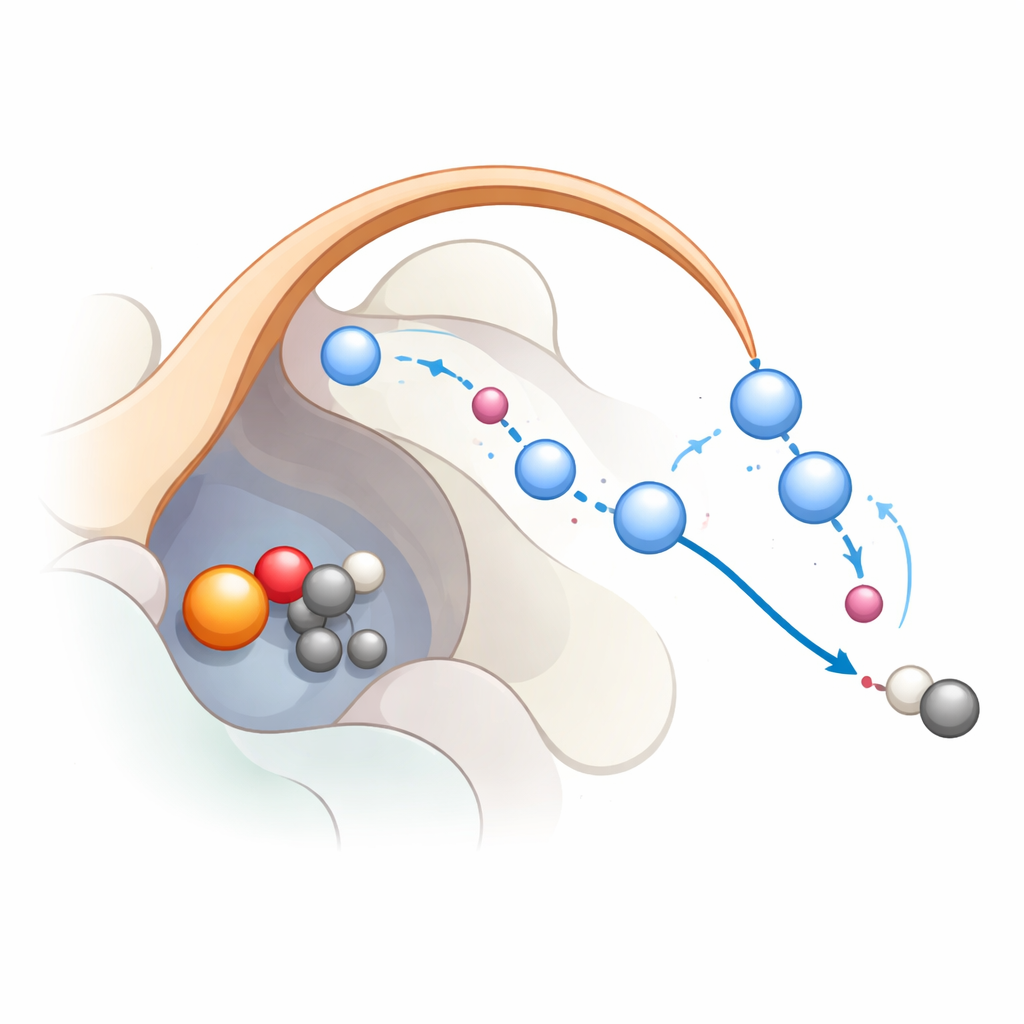
Śledzenie reakcji krok po kroku
Aby zobaczyć, jak ta sieć działa podczas reakcji, badacze połączyli klasyczne symulacje dynamiki molekularnej z zaawansowanymi symulacjami kwantowo‑mechanicznymi. Przyjęli stan, w którym żelazo utworzyło już wysoce reaktywny gatunek żelazo‑okso, powszechny pośrednik w tej rodzinie enzymów. Obliczenia wspierają ścieżkę trzyetapową: najpierw żelazo‑okso zabiera atom wodoru z substratu, tworząc rodnik; następnie następuje przerzut elektronu z powrotem na żelazo, co pomaga substratowi pozbyć się CO2; w trzecim kroku grupa hydroksylowa związana z żelazem zostaje ponownie protonowana, przywracając żelazo do stanu spoczynkowego. Symulacje pokazują, że K191 i łańcuch cząsteczek wody stabilizowanych przez pętelkę klapki tworzą przekaźnik protonowy, który sprawnie przemieszcza protony w ostatnich etapach. Gdy ten przekaźnik jest przerwany, reakcja staje się energetycznie niekorzystna, co dobrze koreluje z danymi mutacyjnymi.
Adaptowalna pętelka kierująca ewolucją enzymu
Ponad samym TraH autorzy porównali spokrewnione enzymy, które mają to samo żelazo‑wiążące jądro, ale wykonują różne zadania chemiczne. Wiele z tych enzymów także ma pętelki przypominające klapki, lecz ich długości, sekwencje i stopnie ruchomości różnią się znacznie. W niektórych pętelka zamyka się mocno nad substratami i tworzy kluczowe wiązania wodorowe; w innych pozostaje otwarta i nie wydaje się uczestniczyć w reakcji. Na podstawie drzew ewolucyjnych i analizy wzorców sekwencji zespół stwierdził, że niektóre reszty pętelki biorące udział w sieci wiązań wodorowych w TraH są silnie konserwowane, podczas gdy inne pozycje zmieniają się w sposób powiązany z preferencjami substratowymi. Sugeruje to, że ewolucja dostraja zarówno sztywność, jak i właściwości chemiczne tych pętelek, by rozpoznawać nowe cząsteczki i kierować reakcje ku określonym wynikom.
Co to oznacza dla przyszłej chemii
Dla osoby niebędącej specjalistą kluczowy wniosek jest taki, że mała elastyczna pętelka na powierzchni enzymu może kontrolować, gdzie protony i cząsteczki wody przemieszczają się wewnątrz, i że ta kontrola może decydować o tym, czy cząsteczka jedynie traci atom wodoru, czy też dodatkowo traci dwutlenek węgla. W TraH rola klapki polega nie tylko na zamknięciu miejsca aktywnego, lecz na złożeniu maleńkiej maszyny transferu protonów z reszt aminokwasowych i uwięzionych cząsteczek wody. Strategia pośrednictwa przez pętelkę różni się od rozwiązania stosowanego przez wiele spokrewnionych enzymów, które używają stałej zasadowej reszty umieszczonej tuż obok odchodzącej grupy. Ujawniając to alternatywne rozwiązanie, praca poszerza nasze rozumienie, jak natura konstruuje wszechstronne katalizatory i dostarcza planu do projektowania enzymów, których ruchome części można przeprogramować, by umożliwić nowe reakcje i nowe bioaktywne cząsteczki.
Cytowanie: Zheng, X., Ge, R., Guo, Z. et al. Lid loop-mediated proton transfer revealed in the Fe/αKG-dependent decarboxylase TraH. Commun Chem 9, 159 (2026). https://doi.org/10.1038/s42004-026-01986-9
Słowa kluczowe: mechanizmy enzymatyczne, niehemowe enzymy żelazowe, transfer protonu, grzybowy metabolizm wtórny, ewolucja enzymów