Clear Sky Science · ja
プロファージがコードする中止感染タンパク質は宿主とプロファージの拡散を維持する
細菌が隠れた同乗者を使ってウイルスと戦う仕組み
細菌はファージと呼ばれるウイルスから常に襲撃を受けているが、多くはプロファージと呼ばれる微小な遺伝的ヒッチハイカー—宿主DNAに組み込まれた休眠ウイルス—のおかげで生き延びている。本研究はそのようなプロファージの一つが、侵入するウイルスを防ぐ強力なセルフデストラクション・スイッチをサルモネラに提供すると同時に、そのプロファージ自身の拡散を維持する仕組みを明らかにする。こうした分子レベルの綱引きを理解することで、病原体がその捕食者に対して一歩先を行く理由が説明される。
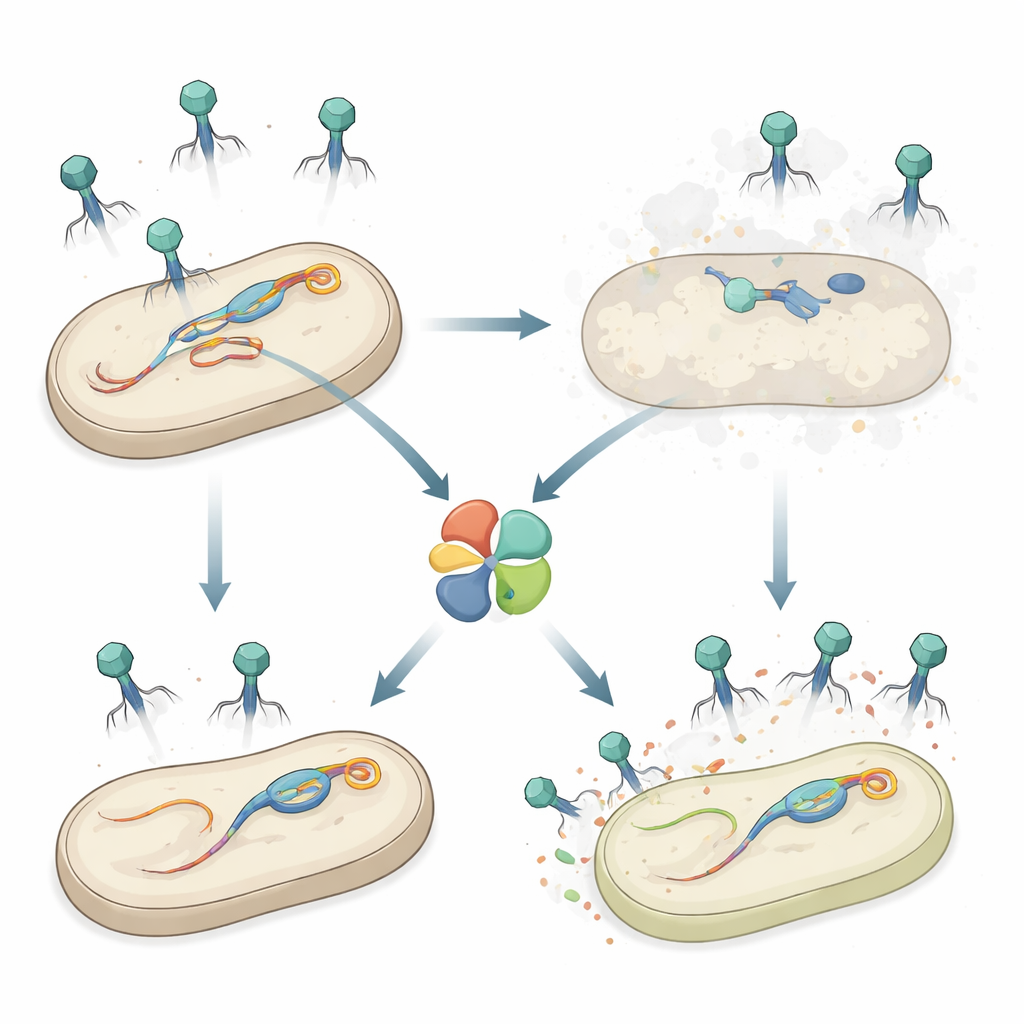
サルモネラの内部にある秘密兵器
研究者らは食中毒原因菌であるSalmonella enterica血清型Typhimuriumを調べた。この菌は多くの細菌と同様に染色体に複数のプロファージを持っている。以前の研究で、Gifsy‑1と呼ばれるプロファージが特定のファージに対する抵抗性に寄与することが示されていた。本研究では、Gifsy‑1上にhepSと名付けられた追加の防御遺伝子を発見し、これが長い尾を持つSiphoviridaeに属する一群のファージに対して細菌の感受性を大幅に低下させることがわかった。hepSを欠失させると、侵入ファージがより多くのプラーク(死細胞の透明な斑点)を形成し、HepSが成功した感染を強く抑えること、また同プロファージ内の他の既知防御機構とは独立して作用することが明らかになった。
攻撃時にのみ作動する自己犠牲システム
HepSはしばしば分子レベルの“最後の抵抗”として機能するタンパク質科に属する:感染を感知すると重要な細胞プロセスを停止させ、細胞とウイルスの両方を死に至らしめる。研究者らはHepSがこのように振る舞う――つまり中止感染システムである――ことを示し、個々の感染細胞を犠牲にすることで集団全体を守ることを示した。構造解析は、HepSが自然に四量体(テトラマー)を形成し、その活性部位は埋もれて不揃いであることを示し、“オフ”状態で宿主にとって無害であることを示唆した。HepSを過剰発現しても細菌の増殖は妨げられず、このタンパク質が正確なトリガーにより作動するまで無害であり続けるという考えを支持した。
ファージ尾端が分子スイッチをどのように反転させるか
HepSを活性化するものを探るため、研究陣はhepSを持つ細菌に多数の異なるファージで挑戦し、感受性の高いウイルスは共通して細菌表面を認識・貫通する尾端タンパク質を持つことを見いだした。HepSを回避して増殖できる稀な“逸脱(エスケープ)”ファージを選択することで、抵抗性は尾端タンパク質Jの短い領域における単一のアミノ酸置換に起因することを突き止めた。細胞内でこのJ断片を発現させるとHepSが毒性を示すが、逸脱型は示さなかった。生化学的実験はHepSがこの小さなJペプチドに直接結合することを確かめた。高分解能結晶構造は、ペプチドがHepS四量体の各サブユニットに結合すると複合体全体が再配列し、活性部位が開いて整列し、HepSが強力なRNA切断酵素へと変換されることを示した。
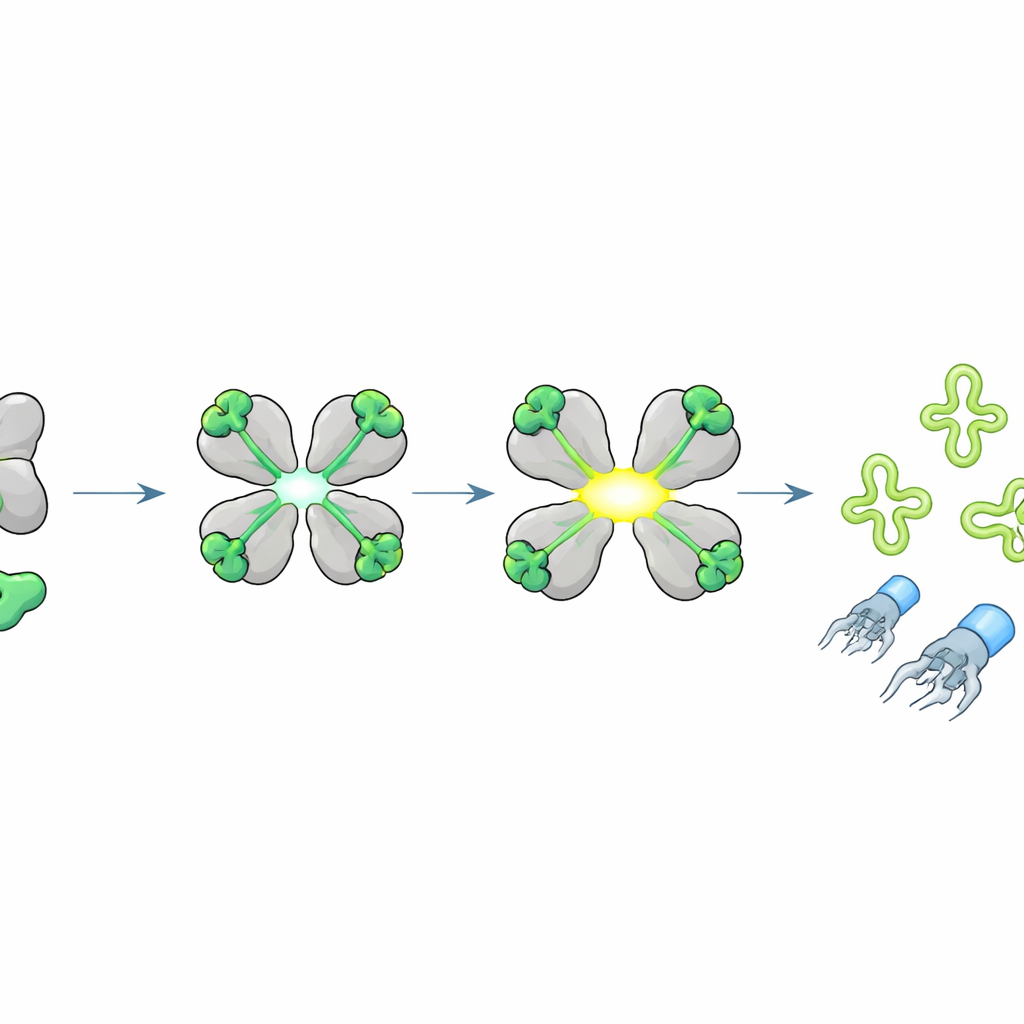
ウイルス産生を停めるために主要なtRNAを切断する
活性化されたHepSは何を切断するのか?研究者らはJペプチドがHepSをオンにすると細胞のRNAが特徴的な断片に分解されることを観察した。タンパク質合成時に遺伝暗号を読み取るアダプターである転移RNA(tRNA)に焦点を当てた専用のシーケンシングを用いると、HepSが特にスレオニン、イソロイシン、セリンに対応するtRNAをアンチコドンループで正確に切断することが明らかになった。構造モデリングは、開いたHepS四量体が標的tRNAを抱え込み、触媒残基をアンチコドンに正確に位置づける様子を示唆した。これらのtRNAを破壊するとタンパク質合成に必要な重要な供給が断たれ、ウイルス性タンパク質の生産が急停止し、完全なファージ粒子の組立が阻害されるが、ウイルスDNAの複製自体は起こり得る、という現象が説明される。
常在プロファージはどのように自家中毒を避けるのか
中心的な謎は、HepSをコードするGifsy‑1自身がなぜ自らの防御で殺されないか、である。その答えもやはり尾端タンパク質にある。サルモネラに存在する三つのGifsyプロファージはいずれもHepSを誘導するファージと近縁のJタンパク質を持つが、活性化するファージが大きなフェニルアラニンを持つ重要部位には、GifsyのJはより穏やかなイソロイシンを持っている。実験は野生型のGifsy JがHepSを活性化しない一方で、フェニルアラニンを入れ替えた改変型は突然HepSを活性化し、hepSが存在する場合に限りGifsy‑1の粒子がほとんど失われることを示した。混合感染では、Gifsy‑1がES18のようなHepSを活性化するプロファージと共存すると、HepSは優先的にES18を無力化し、Gifsy‑1には比較的僅かな影響しか与えないため、HepSをコードするプロファージが新しい粒子を生産して新たな宿主に広がる点で競争優位を得る。
細菌、ウイルス、そして私たちにとっての意義
総じてこの研究は精緻に調律された生存戦略を明らかにした:プロファージはサルモネラ宿主に、関連する侵入ウイルスの特徴を感知する仕掛けタンパク質を供与し、特定のtRNAを切断して細胞のタンパク質合成機構を破壊する。こうした自己犠牲的な一手は侵入者の生活環を止めて広い細菌集団を保護する一方で、プロファージ自身の尾端タンパク質における微妙な差異がそれを回避し、自らの複製を継続させる。本研究は、細菌ゲノムに埋め込まれたウイルスが単に休眠しているだけでなく、他のウイルスとの戦いに能動的に関与し、微生物生態系の中でどの菌株やファージ株が繁栄するかを左右していることを示している。
引用: Sargen, M.R., Antine, S.P., Grabe, G.J. et al. A prophage-encoded abortive infection protein preserves host and prophage spread. Nature 652, 201–208 (2026). https://doi.org/10.1038/s41586-025-10070-6
キーワード: バクテリオファージ防御, プロファージ, 中止感染(abortive infection), tRNAヌクレアーゼ, サルモネラとファージの相互作用