Clear Sky Science · zh
关于RAD51同源蛋白交换如何调控RAD51丝状体形成的结构见解
保护我们的DNA免受日常损伤
你体内的每个细胞都在不断复制和修复其DNA,但在一生中遗传信息却保持出人意料的稳定。当这套修复系统失灵时,突变会积累并可能促成癌症。本文探讨一组辅助蛋白如何协调一项关键的DNA修复步骤,揭示一套分子级的保护机制,它能保持修复高效并避免走入无益的歧路。
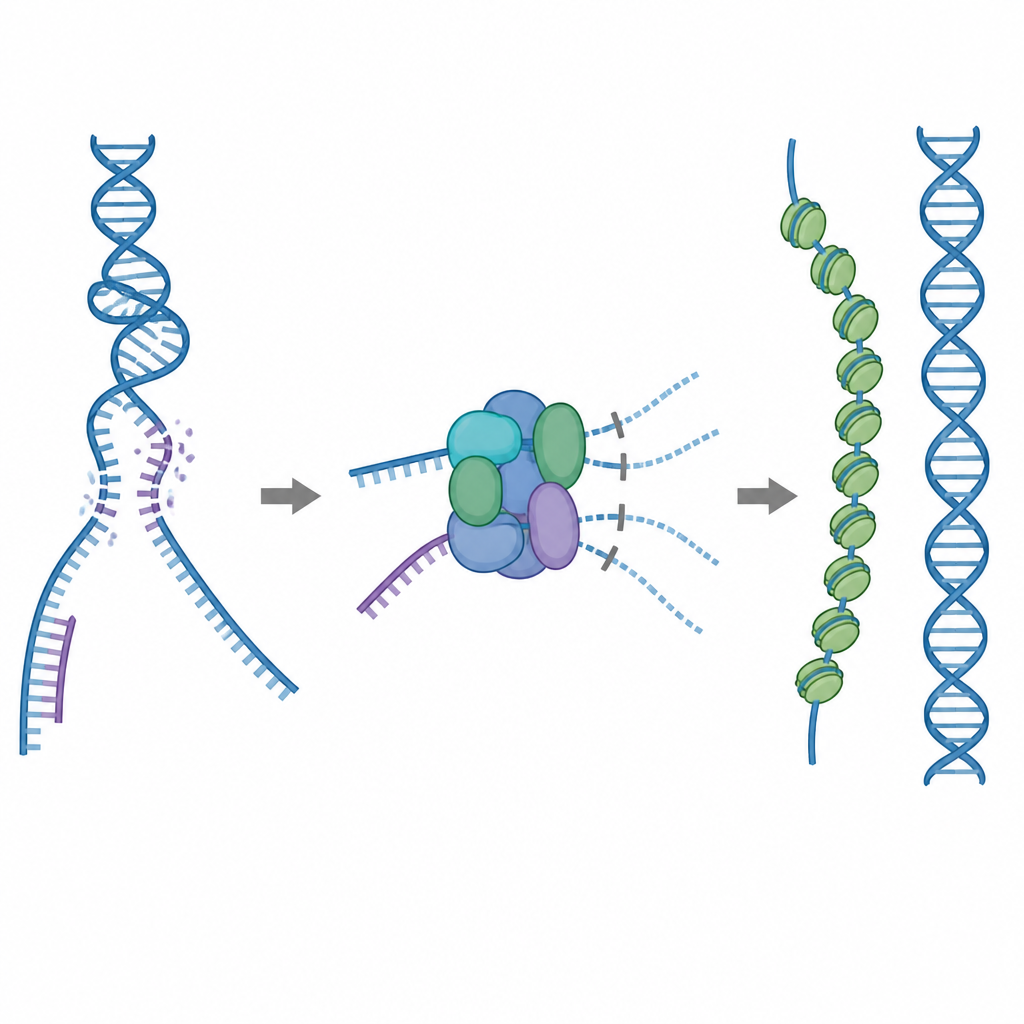
需要谨慎使用的修复工具
细胞的主要修复工具之一是名为RAD51的蛋白,它在破损的单链DNA上形成长丝状体。这些丝状体在完整的DNA拷贝上搜索匹配序列,并协助换链,从而准确修复断裂。但RAD51也可能错误地粘附到不该去的目标上,例如正常的双链DNA或含有RNA–DNA杂合体的结构。如果过多RAD51被困在这些无用的部位,真正的断裂可能得不到修复或被有毒的蛋白–DNA纠缠所干扰。为防止这种情况,细胞利用五种相关的辅助蛋白,即RAD51同源蛋白,但它们的具体功能细节长期不清。
内置的刹车阻止浪费性结合
研究人员利用冷冻电子显微镜可视化了RAD51如何与两种同源蛋白RAD51C和XRCC3联手,形成名为RAD51–X3C的复合物。他们发现八个蛋白单元聚合成环状结构,呈两个四聚体排列。在这种状态下,XRCC3的一段特殊环状区占据了RAD51通常用来抓握DNA的表面,且相邻的RAD51亚基被扭曲错位。生化测试显示,这种“自抑制”复合物仍能相对良好地结合单链DNA,但其与双链DNA或RNA–DNA杂合体的结合能力显著下降。实际上,该复合物起到了刹车的作用,防止RAD51被非生产性的结合位点耗尽,同时又使其保持可用于与单链DNA结合以完成准确修复。
把刹车变成助推器
然而事情并未止步于制动。另两种同源蛋白RAD51D和XRCC2形成了一个名为DX2的伙伴复合物。研究团队表明,DX2可通过停靠在RAD51C暴露的表面上加入RAD51–X3C。这种“同源蛋白交换”将八元环状结构重塑为五元排列,形成RAD51–X3CDX2。在这一新构象中,RAD51用于抓握DNA的面被暴露并正确对齐,形成一段连续、可组装丝状体的表面。重塑后的复合物更紧密地结合单链DNA,帮助RAD51在长链上组装丝状体,并且重要的是,即便这些单链最初被另一种保护蛋白RPA覆盖时,它也能装载RAD51。在体外的链交换反应中,五聚体复合物在RAD51单独存在时强烈受抑制的条件下恢复了其活性。
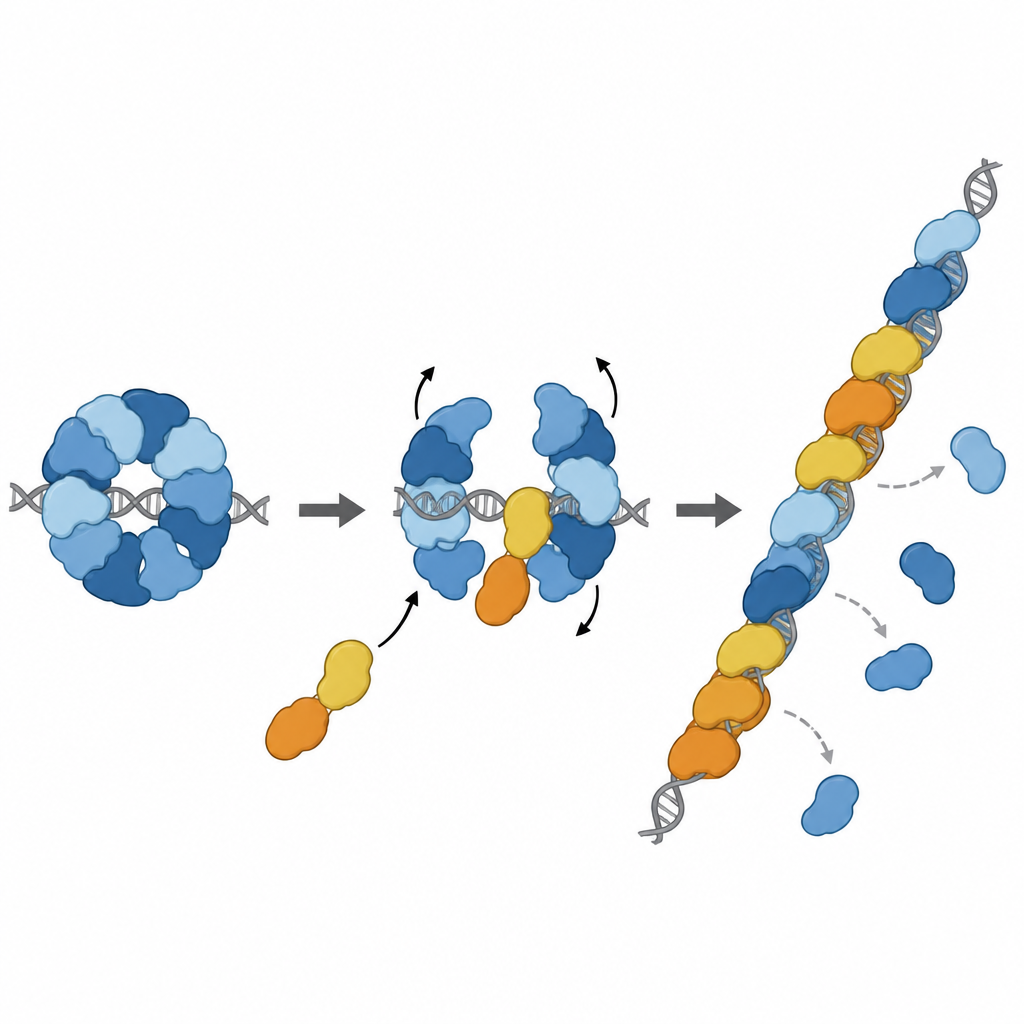
在恰当的位置加入修复行列
进一步的结构研究捕获到了RAD51–X3CDX2复合物实际位于单链DNA上RAD51丝状体末端的情形。在这里,同源蛋白封帽在丝状体的5'端并微妙地改变了附近DNA碱基的保持方式,打破了仅由RAD51构成的丝状体中通常看到的重复三联模式。这提示该复合物既能作为丝状体生长的起始点,又能稳定生长端。细胞实验支持这些结构特征的重要性:破坏XRCC3与RAD51关键接触点的突变削弱了同源重组介导的DNA修复,并损害了停滞复制叉的重新启动——这些过程对维持基因组稳定性至关重要。
对健康与疾病的意义
总体而言,这些发现揭示了针对一个核心DNA修复因子的两步控制系统。首先,RAD51–X3C复合物将RAD51从无益的双链或RNA–DNA区域隔离开,指引其转向断裂的单链DNA。随后,在条件合适时,DX2到来并重塑该装配体为活性的RAD51–X3CDX2单元,从而播种并稳定修复丝状体。因为若干这些同源蛋白的遗传变异与乳腺癌、卵巢癌及其他癌症有关,理解这一分子交接过程有助于更清晰地描绘基因组稳定性通常如何被维持以及其在疾病中如何被破坏的图景。
引用: Rawal, Y., Kwon, Y., Jia, L. et al. Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation. Nat Struct Mol Biol 33, 768–781 (2026). https://doi.org/10.1038/s41594-026-01796-6
关键词: DNA修复, RAD51, 同源重组, 复制应激, 癌症遗传学