Clear Sky Science · pt
Visão estrutural de como a troca de paralogos RAD51 regula a formação de filamentos de RAD51
Protegendo nosso DNA dos danos do dia a dia
Cada célula do seu corpo está constantemente copiando e reparando seu DNA, e ainda assim o roteiro genético permanece notavelmente estável ao longo da vida. Quando esse sistema de reparo falha, mutações podem se acumular e contribuir para o câncer. Este artigo explora como um conjunto de proteínas auxiliares coreografa um passo crucial do reparo do DNA, revelando um mecanismo molecular que mantém o reparo eficiente e evita desvios inúteis.
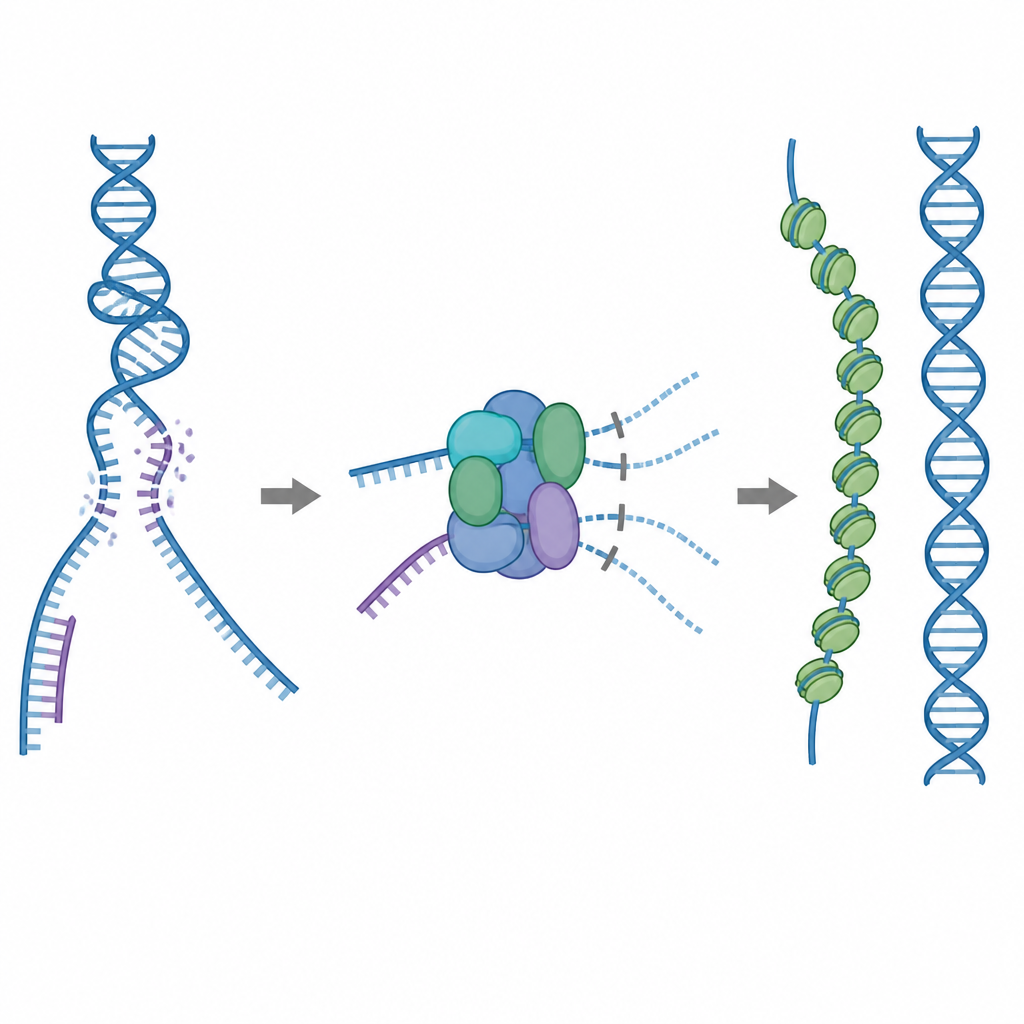
Uma ferramenta de reparo que deve ser usada com cuidado
Uma das principais ferramentas de reparo da célula é uma proteína chamada RAD51, que forma longos filamentos sobre fitas únicas de DNA quebrado. Esses filamentos procuram por uma sequência compatível em uma cópia intacta do DNA e ajudam a trocar as fitas para que a quebra possa ser reparada com precisão. Mas o RAD51 também pode aderir a alvos errados, como DNA de dupla hélice normal ou estruturas que contêm RNA e DNA híbridos. Se muito RAD51 ficar preso nesses locais não produtivos, as quebras verdadeiras podem permanecer sem conserto ou ficar congestionadas com emaranhados tóxicos proteína–DNA. Para evitar isso, as células usam cinco proteínas auxiliares relacionadas, conhecidas como paralogos de RAD51, cujas funções detalhadas têm sido pouco claras.
Um freio embutido que bloqueia ligações inúteis
Usando criomicroscopia eletrônica, os pesquisadores visualizaram como o RAD51 se associa a dois paralogos, RAD51C e XRCC3, para formar um complexo chamado RAD51–X3C. Eles descobriram que oito unidades proteicas se reúnem numa estrutura em forma de anel, organizadas como dois tetrâmeros. Nesse estado, uma alça especial de XRCC3 ocupa fisicamente a superfície do RAD51 que normalmente prende o DNA, e subunidades vizinhas de RAD51 ficam torcidas fora de alinhamento. Testes bioquímicos mostraram que esse complexo “auto-inibido” ainda se liga razoavelmente bem a DNA de fita simples, mas sua capacidade de se prender a DNA de dupla fita ou a híbridos RNA–DNA é fortemente reduzida. Na prática, o complexo atua como um freio que evita que o RAD51 seja absorvido por sítios de ligação não produtivos, ao mesmo tempo em que o deixa livre para interagir com as fitas simples necessárias para um reparo preciso.
Transformando o freio em um impulso
A história não termina no freio. Outros dois paralogos, RAD51D e XRCC2, formam um complexo parceiro chamado DX2. A equipe mostrou que o DX2 pode se juntar ao RAD51–X3C encaixando-se em uma superfície exposta de RAD51C. Essa “troca de paralogos” remodela o anel de oito partes em um arranjo de cinco partes, RAD51–X3CDX2. Nessa nova conformação, a face de RAD51 que prende o DNA fica descoberta e se alinha corretamente, produzindo uma superfície contínua pronta para formar filamentos. O complexo remodelado se liga mais fortemente ao DNA de fita simples, ajuda o RAD51 a montar filamentos em fitas longas e, o mais importante, pode carregar RAD51 mesmo quando essas fitas estão inicialmente recobertas por outra proteína protetora chamada RPA. Em reações laboratoriais de troca de fitas, o complexo pentamérico restabeleceu a atividade do RAD51 em condições em que o RAD51 sozinho estava fortemente inibido.
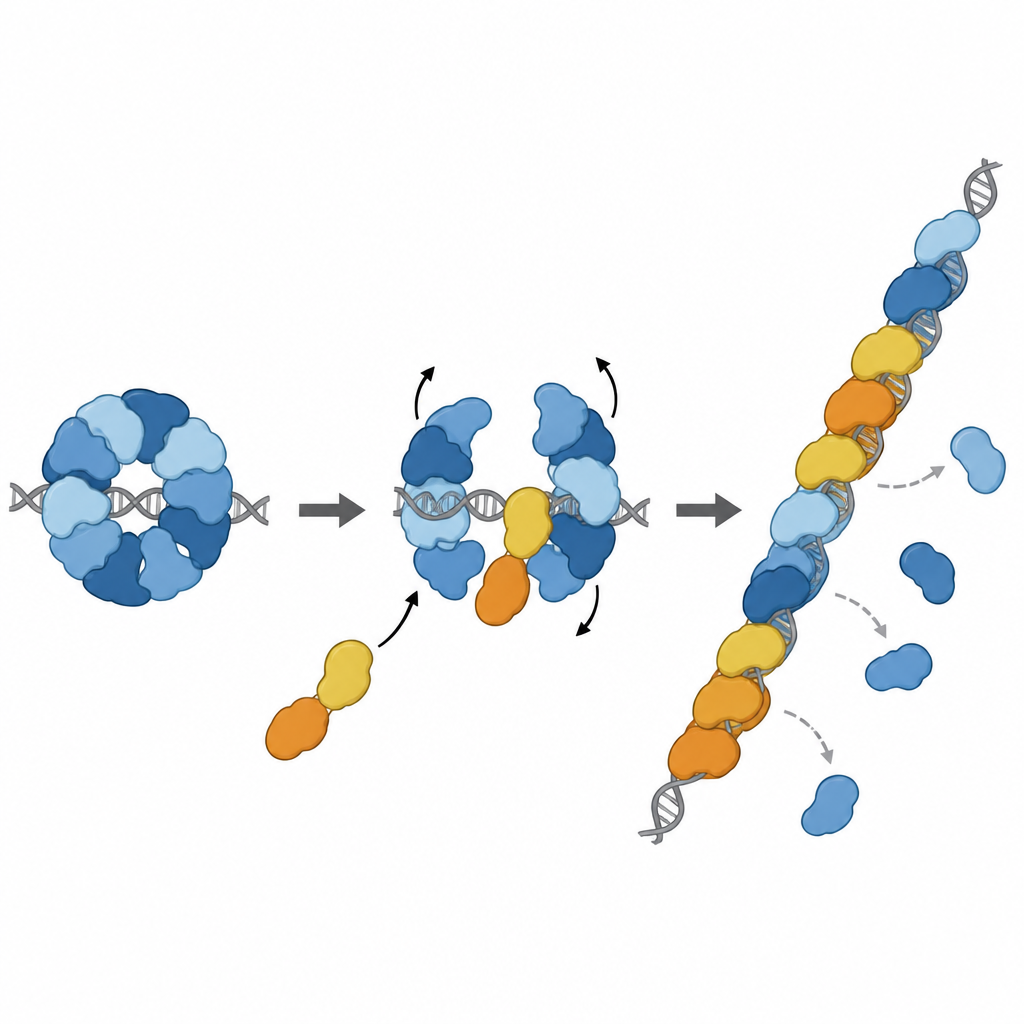
Entrando na fila de reparo no lugar certo
Trabalhos estruturais adicionais capturaram o complexo RAD51–X3CDX2 posicionado na extremidade de um filamento de RAD51 sobre DNA de fita simples. Ali, os paralogos tampam a extremidade 5' do filamento e alteram sutilmente como as bases de DNA próximas são seguradas, rompendo o padrão repetitivo de trios visto em filamentos formados apenas por RAD51. Isso sugere que o complexo tanto semeia o crescimento do filamento quanto estabiliza sua extremidade em expansão. Experimentos em células apoiaram a importância dessas características estruturais: mutações que rompem os pontos de contato chave de XRCC3 com o RAD51 enfraqueceram o reparo por recombinação homóloga e prejudicaram o reinício de forquilhas de replicação estagnadas, processos vitais para manter a estabilidade do genoma.
O que isso significa para saúde e doença
Em conjunto, essas descobertas revelam um sistema de controle em duas etapas para um fator central de reparo do DNA. Primeiro, o complexo RAD51–X3C mantém o RAD51 afastado de regiões não úteis de dupla fita ou híbridos RNA–DNA, direcionando-o para o DNA quebrado de fita simples. Depois, quando as condições são apropriadas, o DX2 chega para remodelar essa montagem em uma unidade ativa RAD51–X3CDX2 que semeia e estabiliza os filamentos de reparo. Como alterações hereditárias em vários desses paralogos estão associadas a cânceres de mama, ovário e outros, compreender essa passagem molecular fornece um quadro mais claro de como a estabilidade do genoma é normalmente preservada e como sua falha pode ocorrer na doença.
Citação: Rawal, Y., Kwon, Y., Jia, L. et al. Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation. Nat Struct Mol Biol 33, 768–781 (2026). https://doi.org/10.1038/s41594-026-01796-6
Palavras-chave: Reparo de DNA, RAD51, Recombinação homóloga, Estresse de replicação, Genética do câncer