Clear Sky Science · it
Prospettiva strutturale su come lo scambio dei paraloghi RAD51 regola la formazione del filamento RAD51
Mantenere il nostro DNA al sicuro dai danni quotidiani
Ogni cellula del tuo corpo copia e ripara costantemente il proprio DNA, eppure il copione genetico rimane straordinariamente stabile per tutta la vita. Quando questo sistema di riparazione fallisce, le mutazioni possono accumularsi e contribuire allo sviluppo del cancro. Questo articolo esplora come un insieme di proteine ausiliarie coreografa un passaggio cruciale della riparazione del DNA, rivelando un meccanismo molecolare che mantiene l’efficacia della riparazione ed evita deviazioni inutili.
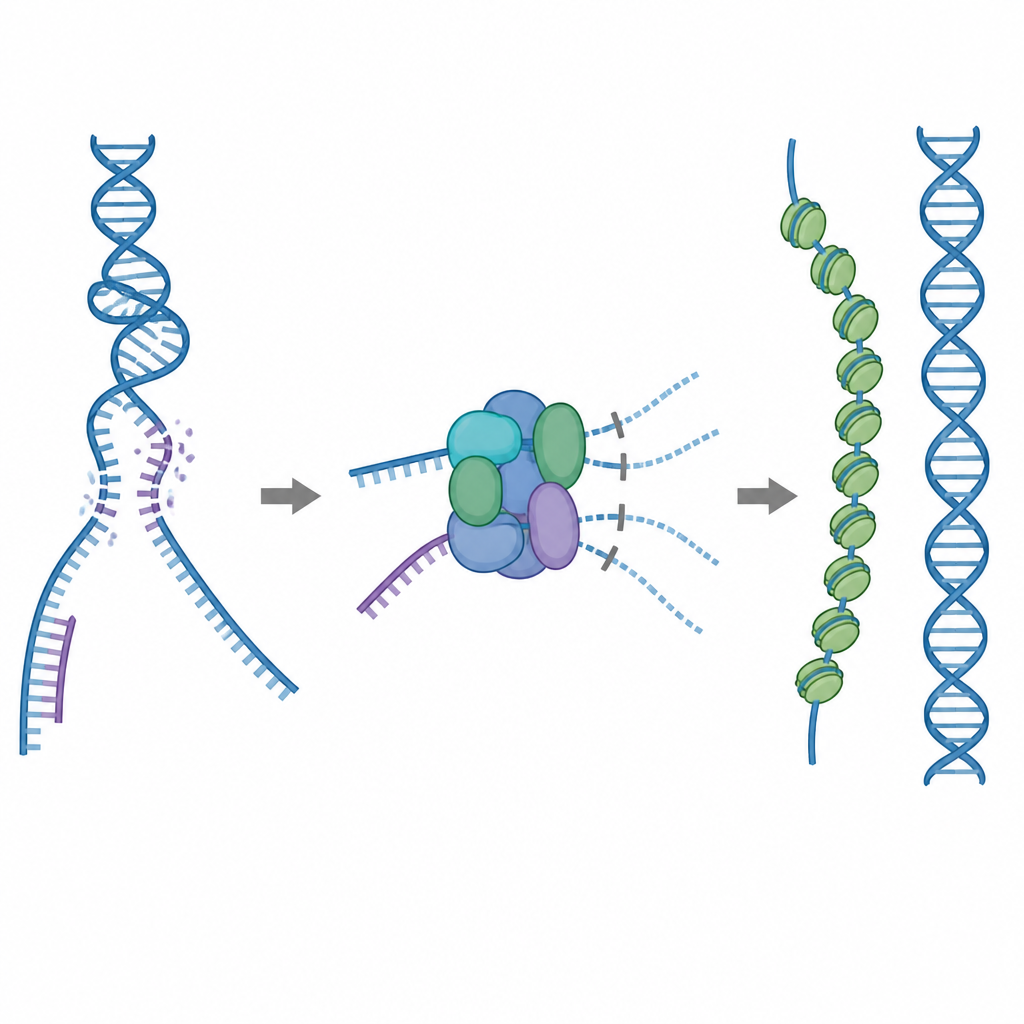
Uno strumento di riparazione da usare con cautela
Uno dei principali strumenti di riparazione della cellula è una proteina chiamata RAD51, che forma lunghi filamenti su filamenti singoli di DNA spezzato. Questi filamenti cercano una sequenza corrispondente su una copia di DNA intatta e aiutano a scambiare i filamenti in modo che la rottura possa essere riparata con accuratezza. Ma RAD51 può anche legarsi a bersagli errati, come il normale DNA a doppio filamento o strutture che contengono sia RNA sia DNA. Se troppo RAD51 è impegnato in questi siti improduttivi, le vere rotture possono rimanere non riparate o intasate da grovigli tossici proteina–DNA. Per prevenire questo, le cellule usano cinque proteine ausiliarie correlate, note come paraloghi di RAD51, i cui ruoli dettagliati non erano chiaramente compresi.
Un freno incorporato che blocca legami improduttivi
Usando la criomicroscopia elettronica, i ricercatori hanno visualizzato come RAD51 si unisca a due paraloghi, RAD51C e XRCC3, per formare un complesso chiamato RAD51–X3C. Hanno scoperto che otto unità proteiche si associano in una struttura ad anello, disposte come due tetrameri. In questo stato, un loop speciale di XRCC3 occupa fisicamente la superficie di RAD51 che normalmente afferra il DNA, e i subunità RAD51 adiacenti risultano torsionate e fuori allineamento. Test biochimici hanno mostrato che questo complesso «autoinibito» si lega ancora abbastanza bene al DNA a filamento singolo, ma la sua capacità di agganciarsi al DNA a doppio filamento o agli ibridi RNA–DNA è fortemente ridotta. In pratica, il complesso agisce come un freno che impedisce a RAD51 di essere assorbito da siti di legame non produttivi, lasciandolo libero di interagire con i filamenti singoli necessari per una riparazione accurata.
Trasformare il freno in un potenziatore
La storia non si ferma al freno. Due altri paraloghi, RAD51D e XRCC2, formano un complesso partner chiamato DX2. Il gruppo ha dimostrato che DX2 può unirsi a RAD51–X3C agganciandosi su una superficie esposta di RAD51C. Questo «scambio di paraloghi» rimodella l’anello a otto parti in un arrangiamento a cinque parti, RAD51–X3CDX2. In questa nuova forma, la faccia di RAD51 che afferra il DNA è scoperta e allineata correttamente, producendo una superficie continua pronta per il filamento. Il complesso rimodellato si lega più saldamente al DNA a filamento singolo, aiuta RAD51 ad assemblare filamenti su filamenti lunghi e, cosa importante, può caricare RAD51 anche quando quei filamenti sono inizialmente ricoperti da un’altra proteina protettiva chiamata RPA. In reazioni biochimiche di scambio di filamenti in laboratorio, il complesso pentamerico ha ripristinato l’attività di RAD51 in condizioni in cui RAD51 da solo era fortemente inibito.
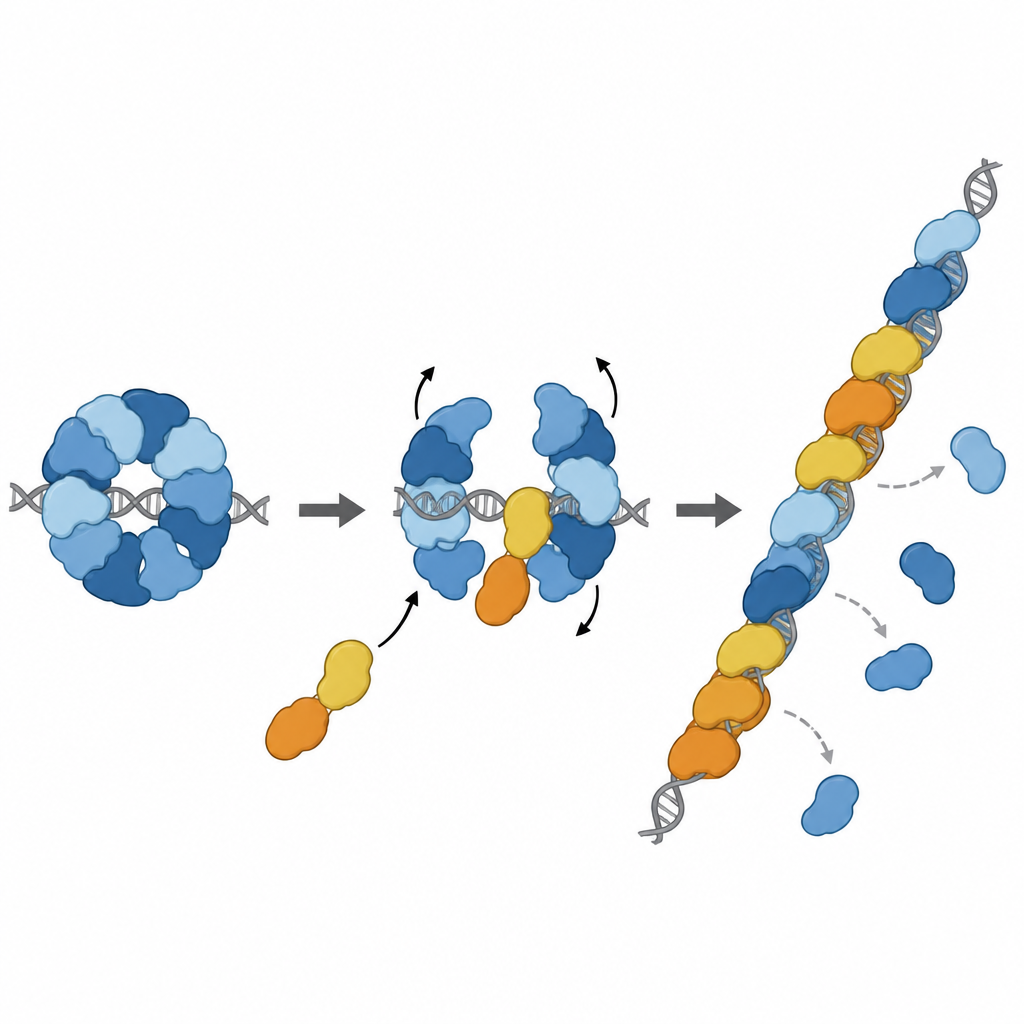
Entrare nella linea di riparazione al momento giusto
Ulteriori analisi strutturali hanno catturato il complesso RAD51–X3CDX2 effettivamente situato all’estremità di un filamento RAD51 su DNA a filamento singolo. Qui, i paraloghi cappano l’estremità a cinque-prime del filamento e alterano sottilmente il modo in cui le basi di DNA vicine sono tenute, interrompendo il solito schema ripetuto a triplette osservato nei filamenti composti solo da RAD51. Ciò suggerisce che il complesso sia sia il nucleo iniziale per la crescita del filamento sia un fattore di stabilizzazione dell’estremità in crescita. Esperimenti in cellule hanno supportato l’importanza di queste caratteristiche strutturali: mutazioni che interrompono i punti di contatto chiave di XRCC3 con RAD51 indeboliscono la riparazione del DNA mediante ricombinazione omologa e compromettono la ripresa delle forcelle di replicazione bloccate, processi vitali per mantenere la stabilità del genoma.
Cosa significa per salute e malattia
Nel complesso, questi risultati rivelano un sistema di controllo in due fasi per un fattore centrale della riparazione del DNA. Innanzitutto, il complesso RAD51–X3C tiene RAD51 lontano da regioni non utili a doppio filamento o da ibridi RNA–DNA, indirizzandolo verso il DNA rotto a filamento singolo. Poi, quando le condizioni sono favorevoli, DX2 arriva per rimodellare questo assemblaggio in un’unità attiva RAD51–X3CDX2 che inocula e stabilizza i filamenti di riparazione. Poiché variazioni ereditarie in diversi di questi paraloghi sono collegate a tumori della mammella, dell’ovaio e altri, comprendere questo passaggio molecolare fornisce un quadro più chiaro di come la stabilità del genoma sia normalmente preservata e di come il suo cedimento possa verificarsi nelle malattie.
Citazione: Rawal, Y., Kwon, Y., Jia, L. et al. Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation. Nat Struct Mol Biol 33, 768–781 (2026). https://doi.org/10.1038/s41594-026-01796-6
Parole chiave: Riparazione del DNA, RAD51, Ricombinazione omologa, Stress di replicazione, Genetica del cancro