Clear Sky Science · tr
RAD51 paralog değişiminin RAD51 filament oluşumunu nasıl düzenlediğine dair yapısal içgörü
Günlük hasarlardan DNA’mızı korumak
Vücudunuzdaki her hücre sürekli olarak DNA’sını kopyalıyor ve onarıyor, yine de genetik metin yaşam boyu boyunca şaşırtıcı derecede sabit kalıyor. Bu onarım sistemi aksadığında mutasyonlar birikebilir ve kansere katkıda bulunabilir. Bu makale, bir dizi yardımcı proteinin kritik bir DNA onarım adımını nasıl düzenlediğini inceliyor ve onarımın verimli kalmasını sağlayan, gereksiz sapmalardan kaçınan moleküler bir emniyet mekanizmasını ortaya koyuyor.
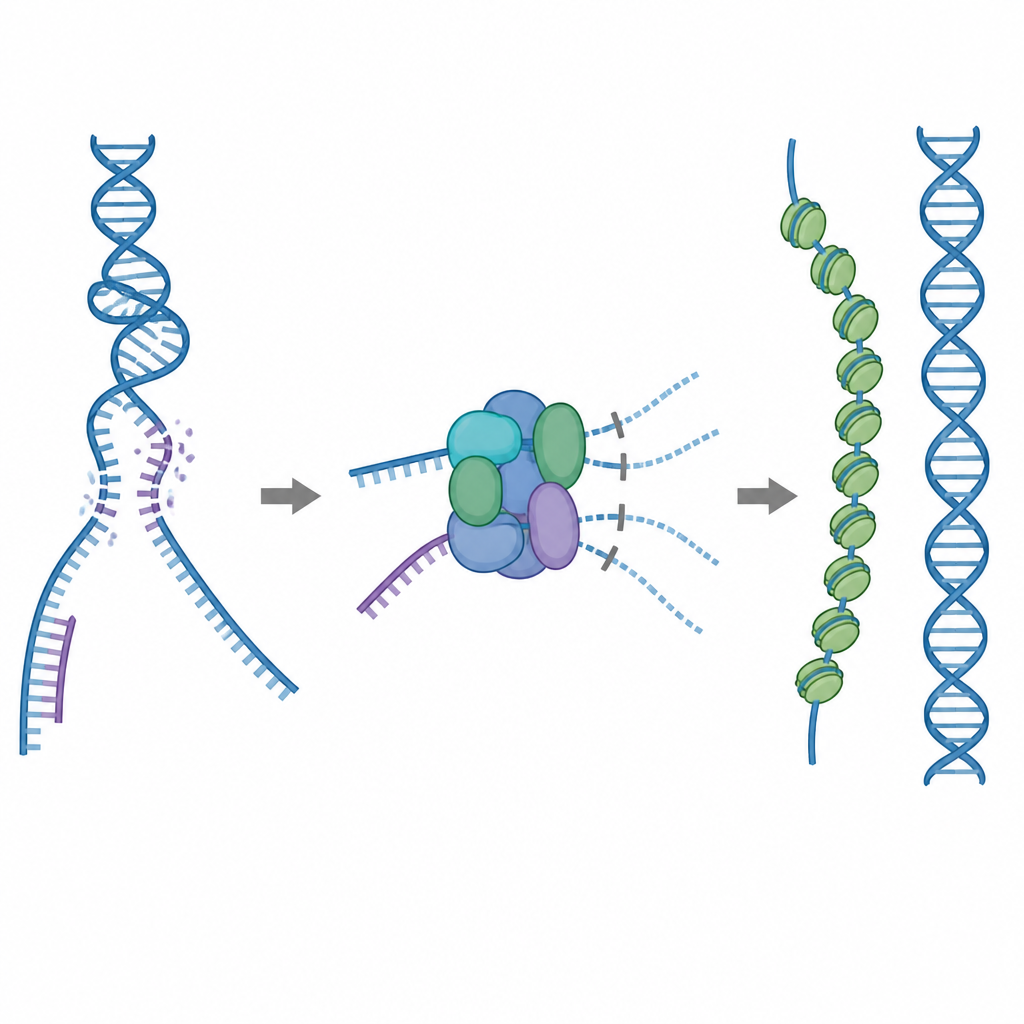
Dikkatle kullanılması gereken bir onarım aracı
Hücrenin ana onarım araçlarından biri RAD51 adlı proteindir; bu protein kırık tek iplikli DNA üzerinde uzun filamentler oluşturur. Bu filamentler sağlam bir DNA kopyasında eşleşen diziyi arar ve kırığın doğru şekilde onarılabilmesi için zincir değişimini kolaylaştırır. Ancak RAD51 yanlış hedeflere, örneğin normal çift sarmallı DNA’ya veya RNA–DNA hibrit yapılarına da yapışabilir. Çok fazla RAD51 bu işe yaramayan noktalara bağlanırsa gerçek kırıklar onarılmadan kalabilir veya toksik protein–DNA düğümleriyle dolabilir. Bunu önlemek için hücreler RAD51 paralogları olarak bilinen beş ilişkili yardımcı proteini kullanır; bu proteinlerin ayrıntılı görevleri ise belirsizdi.
Atıl bağlanmayı engelleyen yerleşik bir fren
Kryo elektron mikroskobu kullanarak araştırmacılar, RAD51’in iki paralogu RAD51C ve XRCC3 ile nasıl birleşip RAD51–X3C adlı bir kompleks oluşturduğunu görselleştirdiler. Sekiz protein biriminin halka benzeri bir yapı oluşturmak üzere iki tetramer olarak düzenlendiğini buldular. Bu durumda XRCC3’ten gelen özel bir döngü, normalde DNA’yı kavrayan RAD51 yüzeyini fiziksel olarak işgal eder ve komşu RAD51 alt birimleri hizalanmadan bükülür. Biyokimyasal testler, bu "oto-inkine edilmiş" kompleksin tek iplikli DNA’ya hâlâ makul düzeyde bağlandığını, ancak çift sarmallı DNA veya RNA–DNA hibritlerine tutunma yeteneğinin keskin bir şekilde azaldığını gösterdi. Etkili bir şekilde kompleks, RAD51’in üretken olmayan bağlanma bölgelerine kapılıp gitmesini engelleyen bir fren görevi görürken, doğru onarım için gerekli tek ipliklerle etkileşime girmesine izin verir.
Freni güçlendiriciye çevirme
Hikaye sadece frenle bitmiyor. Diğer iki paralog, RAD51D ve XRCC2, DX2 adı verilen bir ortak kompleks oluşturur. Ekip, DX2’nin RAD51–X3C’ye RAD51C’nin açıkta kalan bir yüzeyine kenetlenerek katılabileceğini gösterdi. Bu "paralog değişimi" sekiz parçalı halkayı beş parçalı bir düzen olan RAD51–X3CDX2’ye yeniden şekillendirir. Bu yeni yapıda RAD51’in DNA’yı kavrayan yüzeyi açığa çıkar ve doğru şekilde hizalanır; böylece sürekli, filament oluşturulmaya hazır bir yüzey oluşur. Yeniden modellenmiş kompleks tek iplikli DNA’ya daha sıkı bağlanır, RAD51’in uzun zincirler üzerinde filament oluşturmasına yardımcı olur ve önemli olarak, bu zincirler başlangıçta başka bir koruyucu protein olan RPA ile kaplı olsa bile RAD51’i yükleyebilir. Laboratuvar strand-değişim reaksiyonlarında, pentamerik kompleks RAD51 tek başına güçlü şekilde inhibe edildiği koşullarda RAD51’in aktivitesini geri getirdi.
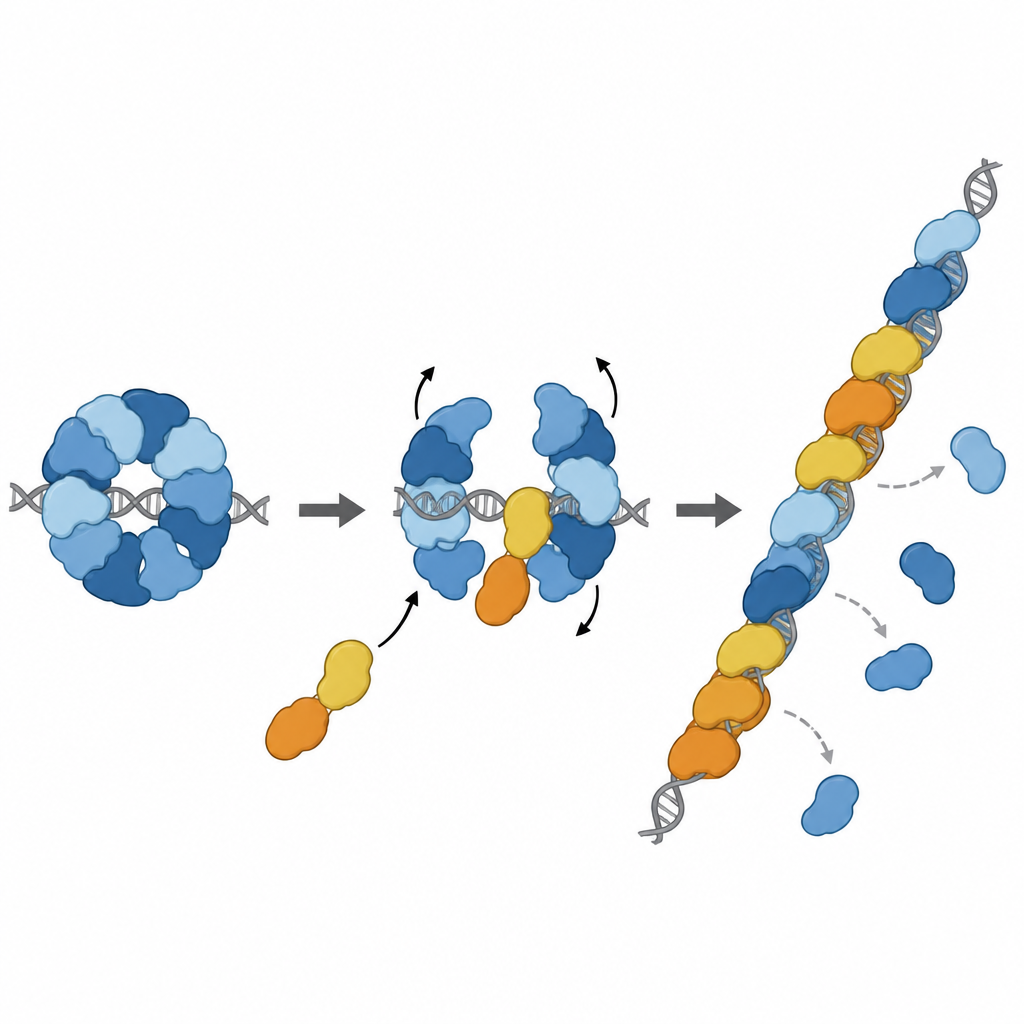
Onarım sırasına tam da doğru yerde katılmak
Daha fazla yapısal çalışma, RAD51–X3CDX2 kompleksinin tek iplikli DNA üzerindeki bir RAD51 filamentinin ucunda gerçekten oturduğunu yakaladı. Burada paraloglar filamentin beş-prime ucunu kapatır ve yakınlardaki DNA bazlarının tutulma şeklini hafifçe değiştirerek RAD51 yalnız filamentlerinde görülen tekrarlayan üçlü düzeni bozar. Bu bulgu, kompleksin hem filament büyümesini başlattığını hem de büyüyen ucu stabilize ettiğini öne sürer. Hücre temelli deneyler bu yapısal özelliklerin önemini destekledi: XRCC3’ün RAD51 ile yaptığı kritik temas noktalarını bozan mutasyonlar homolog rekombinasyonla DNA onarımını zayıflattı ve duraklamış DNA replikasyon çatallarının yeniden başlatılmasını bozdu; bu süreçler genom stabilitesinin korunması için hayati öneme sahiptir.
Bu bulguların sağlık ve hastalık açısından anlamı
Bu bulgular birlikte merkezi bir DNA onarım faktörü için iki aşamalı bir kontrol sistemini ortaya koyuyor. Önce RAD51–X3C kompleksi RAD51’i işe yaramayan çift sarmallı veya RNA–DNA bölgelerinden uzak tutar ve onu kırık, tek iplikli DNA’ya yönlendirir. Ardından, koşullar uygun olduğunda DX2 gelip bu düzeni aktif bir RAD51–X3CDX2 ünitesine yeniden şekillendirir; bu ünite onarım filamentlerini başlatır ve stabilize eder. Bu paralogların birkaçında kalıtsal değişikliklerin meme, yumurtalık ve diğer kanserlerle ilişkilendirilmiş olması nedeniyle bu moleküler el değişimini anlamak, genom stabilitesinin normalde nasıl korunduğu ve hastalıkta bunun nasıl bozulabileceğine dair daha net bir tablo sağlar.
Atıf: Rawal, Y., Kwon, Y., Jia, L. et al. Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation. Nat Struct Mol Biol 33, 768–781 (2026). https://doi.org/10.1038/s41594-026-01796-6
Anahtar kelimeler: DNA onarımı, RAD51, homolog rekombinasyon, replikasyon stresi, kanser genetiği