Clear Sky Science · fr
Aperçu structural de la manière dont l’échange de paralogues RAD51 régule la formation des filaments RAD51
Protéger notre ADN contre les dommages du quotidien
Chaque cellule de votre corps copie et répare en permanence son ADN, et pourtant le script génétique reste remarquablement stable au fil de la vie. Lorsque ce système de réparation déraille, des mutations peuvent s’accumuler et contribuer au développement du cancer. Cet article examine comment un ensemble de protéines auxiliaires orchestre une étape cruciale de la réparation de l’ADN, révélant un mécanisme moléculaire qui maintient l’efficacité de la réparation et évite les détours inutiles.
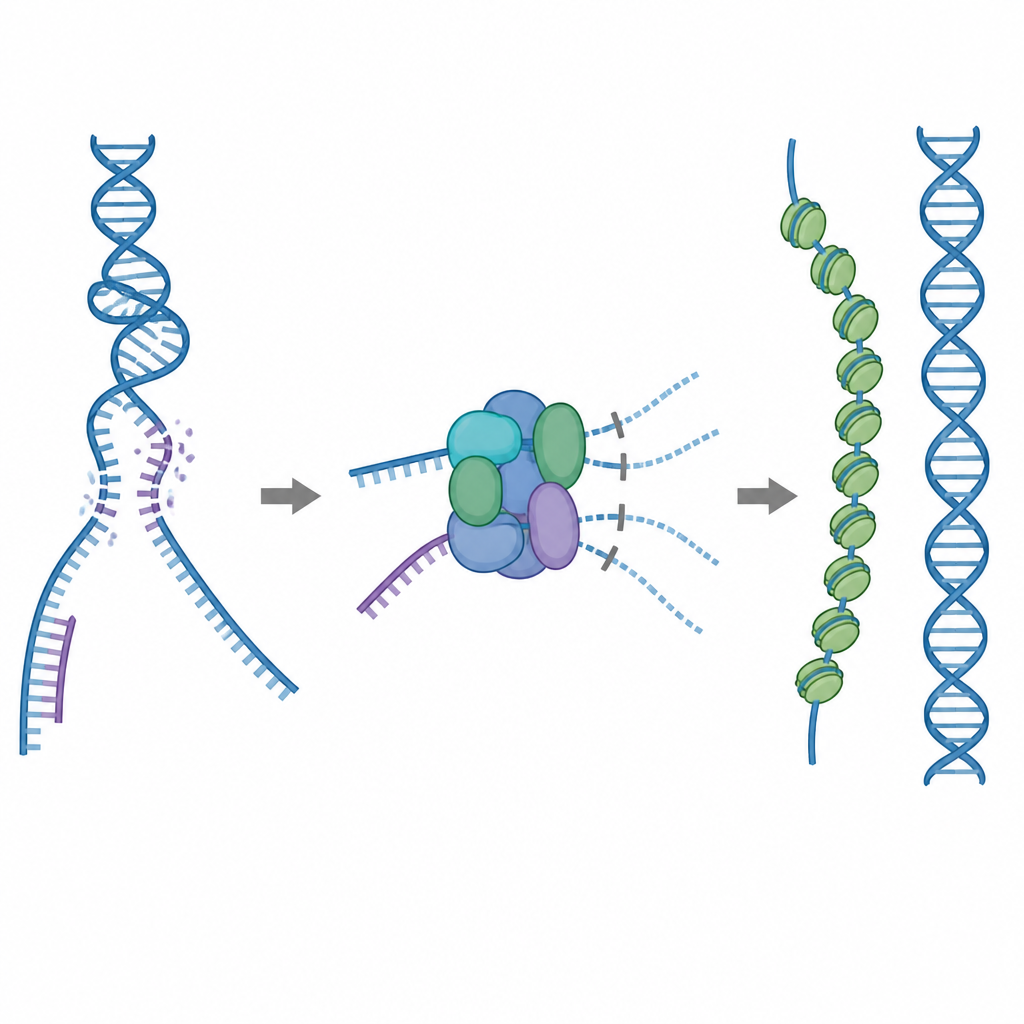
Un outil de réparation à utiliser avec précaution
Un des principaux outils de réparation de la cellule est une protéine appelée RAD51, qui forme de longs filaments sur les brins simples d’ADN cassé. Ces filaments recherchent une séquence correspondante sur une copie intacte de l’ADN et facilitent l’échange de brins pour que la cassure soit réparée avec précision. Mais RAD51 peut aussi se lier à de mauvaises cibles, comme l’ADN double brin normal ou des structures hybrides ARN–ADN. Si trop de RAD51 est immobilisé dans ces sites non productifs, les véritables cassures peuvent rester non réparées ou encombrées par des enchevêtrements protéine–ADN toxiques. Pour l’empêcher, les cellules utilisent cinq protéines auxiliaires apparentées, appelées paralogues de RAD51, dont les rôles précis restaient flous.
Un frein intégré qui empêche les liaisons inutiles
Grâce à la cryo‑microscopie électronique, les chercheurs ont visualisé comment RAD51 s’assemble avec deux paralogues, RAD51C et XRCC3, pour former un complexe appelé RAD51–X3C. Ils ont constaté que huit unités protéiques se réunissent en une structure en anneau, organisée en deux tétramères. Dans cet état, une boucle spécifique de XRCC3 occupe physiquement la surface de RAD51 qui se lie normalement à l’ADN, et les sous‑unités RAD51 voisines sont décalées hors d’alignement. Des essais biochimiques ont montré que ce complexe « autoinhibé » se lie encore assez bien à l’ADN simple brin, mais que sa capacité à se fixer à l’ADN double brin ou aux hybrides ARN–ADN est fortement réduite. En pratique, le complexe agit comme un frein qui empêche RAD51 d’être aspiré par des sites de liaison non productifs tout en le laissant libre de reconnaître les brins simples nécessaires à une réparation fidèle.
Transformer le frein en amplificateur
L’histoire ne s’arrête pas au freinage. Deux autres paralogues, RAD51D et XRCC2, forment un complexe partenaire nommé DX2. L’équipe a montré que DX2 peut s’intégrer à RAD51–X3C en s’amarrant sur une surface exposée de RAD51C. Cet « échange de paralogues » remodèle l’anneau huit‑parties en une organisation à cinq composants, RAD51–X3CDX2. Dans cette nouvelle conformation, la face de RAD51 qui saisit l’ADN est dégagée et correctement alignée, produisant une surface continue prête à former un filament. Le complexe remanié se lie plus fortement à l’ADN simple brin, aide RAD51 à assembler des filaments sur de longs brins et, fait important, peut charger RAD51 même lorsque ces brins sont initialement recouverts par une autre protéine protectrice appelée RPA. Dans des réactions d’échange de brins en laboratoire, le complexe pentamérique a restauré l’activité de RAD51 là où RAD51 seul était fortement inhibé.
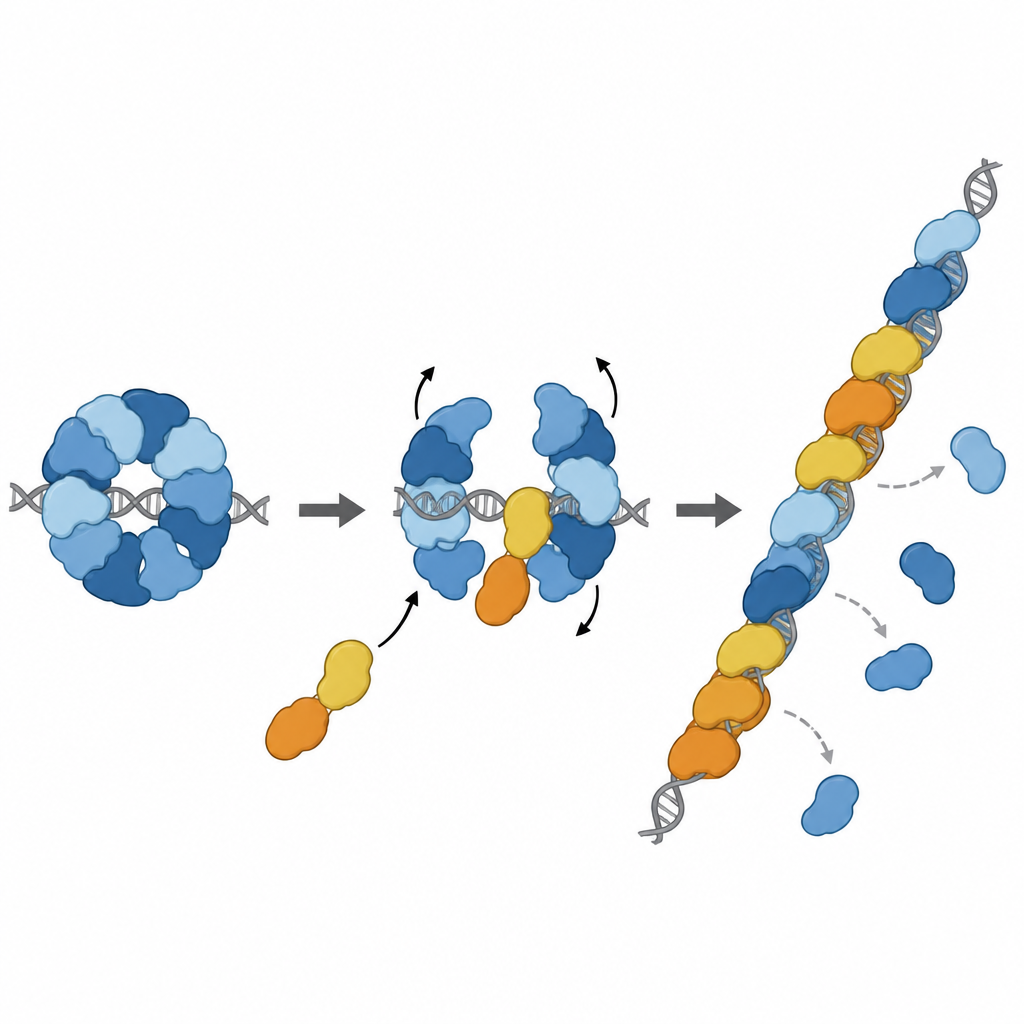
S’engager sur la ligne de réparation au bon endroit
Des travaux structuraux supplémentaires ont capturé le complexe RAD51–X3CDX2 posé à l’extrémité d’un filament RAD51 sur de l’ADN simple brin. Ici, les paralogues coiffent l’extrémité 5′ du filament et modifient subtilement la façon dont les bases d’ADN voisines sont maintenues, rompant le motif répétitif en triplets habituellement observé dans les filaments composés uniquement de RAD51. Cela suggère que le complexe amorce la croissance du filament et stabilise l’extrémité en expansion. Des expériences cellulaires ont confirmé l’importance de ces caractéristiques structurales : des mutations perturbant les points de contact clés de XRCC3 avec RAD51 affaiblissaient la réparation de l’ADN par recombinaison homologue et entravaient le redémarrage des fourches de réplication bloquées, processus essentiels au maintien de la stabilité du génome.
Implications pour la santé et la maladie
Ensemble, ces résultats révèlent un système de contrôle en deux étapes pour un facteur central de la réparation de l’ADN. D’abord, le complexe RAD51–X3C éloigne RAD51 des régions non utiles double brin ou ARN–ADN, le dirigeant vers l’ADN simple brin lésé. Puis, lorsque les conditions sont réunies, DX2 arrive pour remodeler cet assemblage en une unité active RAD51–X3CDX2 qui amorce et stabilise les filaments de réparation. Parce que des altérations héréditaires dans plusieurs de ces paralogues sont liées au cancer du sein, de l’ovaire et à d’autres cancers, comprendre ce relais moléculaire offre une image plus nette de la manière dont la stabilité du génome est normalement préservée et de la façon dont sa défaillance peut conduire à la maladie.
Citation: Rawal, Y., Kwon, Y., Jia, L. et al. Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation. Nat Struct Mol Biol 33, 768–781 (2026). https://doi.org/10.1038/s41594-026-01796-6
Mots-clés: réparation de l’ADN, RAD51, recombinaison homologue, stress de réplication, génétique du cancer