Clear Sky Science · en
Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation
Keeping our DNA safe from everyday damage
Every cell in your body is constantly copying and repairing its DNA, yet the genetic script remains remarkably stable over a lifetime. When this repair system falters, mutations can build up and contribute to cancer. This article explores how a set of helper proteins choreograph a crucial DNA repair step, revealing a molecular safeguard that keeps repair efficient and avoids wasteful detours.
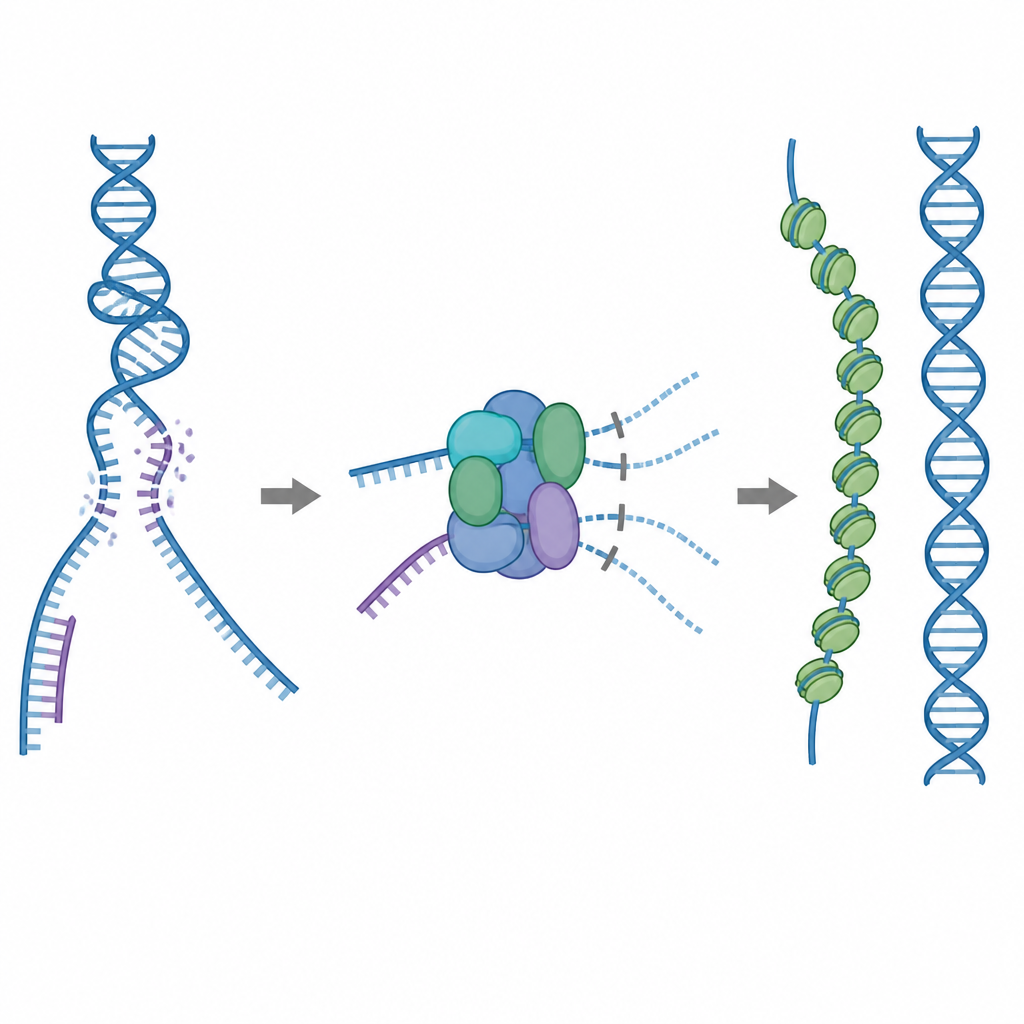
A repair tool that must be used with care
One of the cell’s main repair tools is a protein called RAD51, which forms long filaments on single strands of broken DNA. These filaments search for a matching sequence on an intact DNA copy and help swap strands so the break can be fixed accurately. But RAD51 can also stick to the wrong targets, such as normal double-stranded DNA or structures that contain both RNA and DNA. If too much RAD51 is tied up in these unhelpful spots, the true breaks may be left unrepaired or cluttered with toxic protein–DNA tangles. To prevent this, cells use five related helper proteins, known as RAD51 paralogs, whose detailed jobs have been unclear.
A built in brake that blocks wasteful binding
Using cryo electron microscopy, the researchers visualized how RAD51 teams up with two paralogs, RAD51C and XRCC3, to form a complex called RAD51–X3C. They found that eight protein units come together in a ring-like structure, arranged as two tetramers. In this state, a special loop from XRCC3 physically occupies the surface of RAD51 that normally grips DNA, and neighboring RAD51 subunits are twisted out of alignment. Biochemical tests showed that this "autoinhibited" complex still binds single-stranded DNA reasonably well, but its ability to latch onto double-stranded DNA or RNA–DNA hybrids is sharply reduced. In effect, the complex acts as a brake that keeps RAD51 from being soaked up by nonproductive binding sites while leaving it free to engage the single strands needed for accurate repair.
Turning the brake into a booster
The story does not end with braking. Two other paralogs, RAD51D and XRCC2, form a partner complex called DX2. The team showed that DX2 can join RAD51–X3C by docking onto an exposed surface of RAD51C. This "paralog exchange" remodels the eight-part ring into a five-part arrangement, RAD51–X3CDX2. In this new shape, RAD51’s DNA-gripping face is uncovered and lines up properly, producing a continuous, filament-ready surface. The remodeled complex binds single-stranded DNA more tightly, helps RAD51 assemble filaments on long strands, and, importantly, can load RAD51 even when those strands are initially coated by another protective protein called RPA. In laboratory strand-exchange reactions, the pentameric complex restored RAD51’s activity under conditions where RAD51 alone was strongly inhibited.
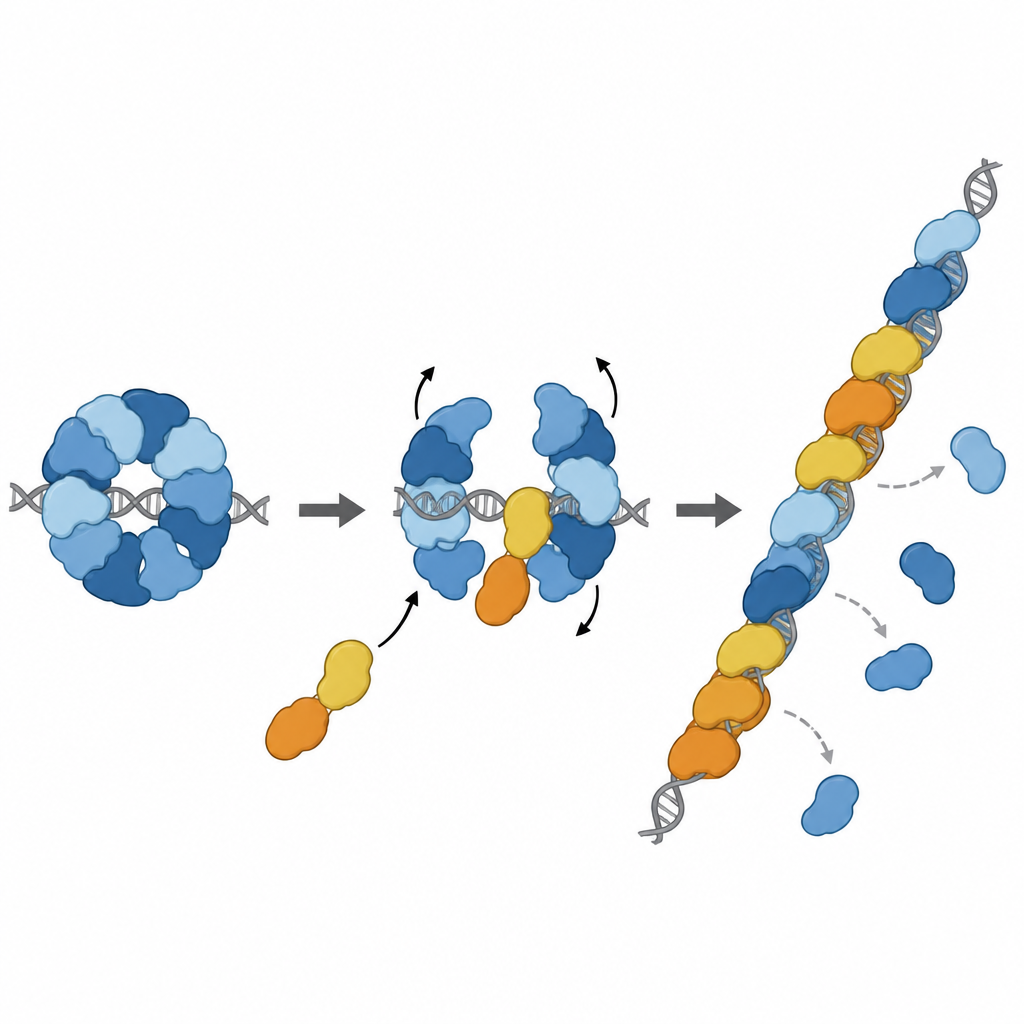
Joining the repair line at just the right place
Further structural work captured the RAD51–X3CDX2 complex actually sitting at the end of a RAD51 filament on single-stranded DNA. Here, the paralogs cap the five-prime end of the filament and subtly alter how nearby DNA bases are held, breaking the usual repeating triplet pattern seen in RAD51-only filaments. This suggests that the complex both seeds filament growth and stabilizes the growing end. Cell-based experiments supported the importance of these structural features: mutations that disrupt XRCC3’s key contact points with RAD51 weakened DNA repair by homologous recombination and impaired the restart of stalled DNA replication forks, processes that are vital for maintaining genome stability.
What this means for health and disease
Together, these findings reveal a two-step control system for a central DNA repair factor. First, the RAD51–X3C complex keeps RAD51 away from unhelpful double-stranded or RNA–DNA regions, steering it toward broken, single-stranded DNA. Then, when conditions are right, DX2 arrives to remodel this assembly into an active RAD51–X3CDX2 unit that seeds and stabilizes repair filaments. Because inherited changes in several of these paralogs are linked to breast, ovarian and other cancers, understanding this molecular handoff provides a clearer picture of how genome stability is normally preserved and how its breakdown may occur in disease.
Citation: Rawal, Y., Kwon, Y., Jia, L. et al. Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation. Nat Struct Mol Biol 33, 768–781 (2026). https://doi.org/10.1038/s41594-026-01796-6
Keywords: DNA repair, RAD51, homologous recombination, replication stress, cancer genetics