Clear Sky Science · ru
Структурное понимание того, как обмен паралогов RAD51 регулирует формирование филамента RAD51
Защита нашей ДНК от повседневных повреждений
Каждая клетка вашего организма постоянно копирует и ремонтирует свою ДНК, но за всю жизнь генетическая информация остаётся удивительно стабильной. Когда эта система репарации даёт сбой, мутации накапливаются и могут способствовать развитию рака. В этой статье рассказывается о том, как набор вспомогательных белков координирует ключевой шаг ремонта ДНК, раскрывая молекулярный предохранитель, который делает репарацию эффективной и избегает бесполезных отклонений.
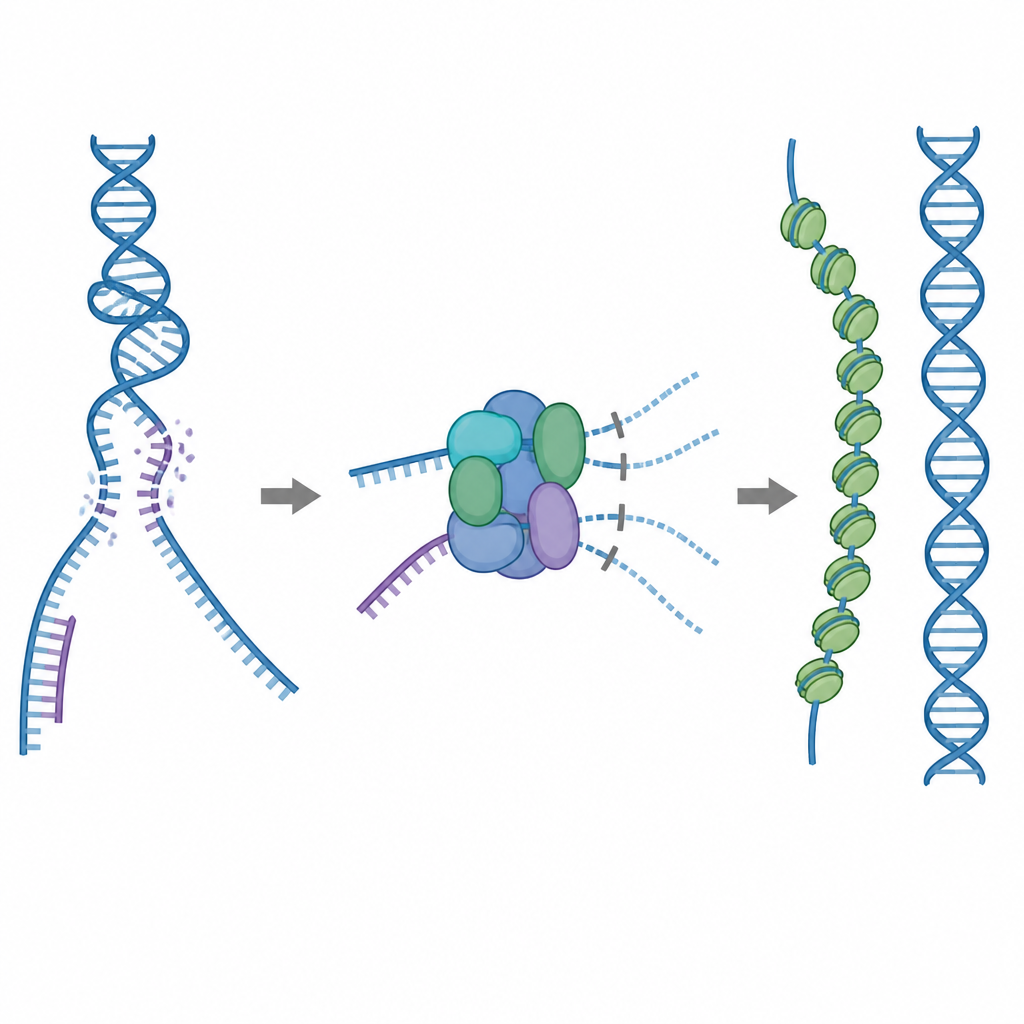
Инструмент ремонта, требующий осторожного обращения
Одним из основных инструментов клетки для ремонта является белок RAD51, который образует длинные филаменты на одноцепочечной разорванной ДНК. Эти филаменты ищут совпадающую последовательность на неповреждённой копии ДНК и помогают обменять цепи, чтобы разрыв можно было исправить с точностью. Но RAD51 также может прикрепляться не к тем целям — например, к обычной двухцепочечной ДНК или к структурам, содержащим гибриды РНК–ДНК. Если слишком много RAD51 окажется связанным в таких неполезных участках, истинные разрывы могут остаться неремонтированными или облиться токсичными белково‑ДНК связями. Чтобы этого избежать, клетки используют пять родственных вспомогательных белков, известных как паралоги RAD51, роли которых ранее были не до конца ясны.
Встроенный тормоз, блокирующий бесполезное связывание
С помощью криоэлектронной микроскопии исследователи визуализировали, как RAD51 взаимодействует с двумя паралогами, RAD51C и XRCC3, образуя комплекс, названный RAD51–X3C. Они обнаружили, что восемь белковых субъединиц собираются в кольцеобразную структуру, организованную как два тетрамера. В этом состоянии специальная петля от XRCC3 физически занимает поверхность RAD51, которая обычно захватывает ДНК, а соседние субъединицы RAD51 вывернуты из выравнивания. Биохимические тесты показали, что этот «аутокинетически подавленный» комплекс по‑прежнему довольно хорошо связывает одноцепочечную ДНК, но его способность прикрепляться к двухцепочечной ДНК или гибридам РНК–ДНК резко снижена. По сути, комплекс действует как тормоз, не давая RAD51 расходоваться на непродуктивные сайты связывания, оставляя его доступным для работы с одноцепочечными участками, необходимыми для точного ремонта.
Преобразование тормоза в усилитель
История не заканчивается на торможении. Два других паралога, RAD51D и XRCC2, образуют партнёрский комплекс, названный DX2. Авторы показали, что DX2 может присоединяться к RAD51–X3C, докируясь к оголённой поверхности RAD51C. Этот «обмен паралогов» перестраивает восьмизвенную кольцевую структуру в пятизвенную конфигурацию RAD51–X3CDX2. В новом виде лицевая поверхность RAD51, захватывающая ДНК, освобождается и выравнивается, образуя непрерывную поверхность, готовую для формирования филамента. Перестроенный комплекс сильнее связывает одноцепочечную ДНК, способствует сборке филаментов RAD51 на длинных нитях и, что важно, способен загружать RAD51 даже когда эти нити первоначально покрыты другим защитным белком, RPA. В лабораторных реакциях обмена цепями пентамерный комплекс восстановил активность RAD51 в условиях, где один RAD51 был сильно ингибирован.
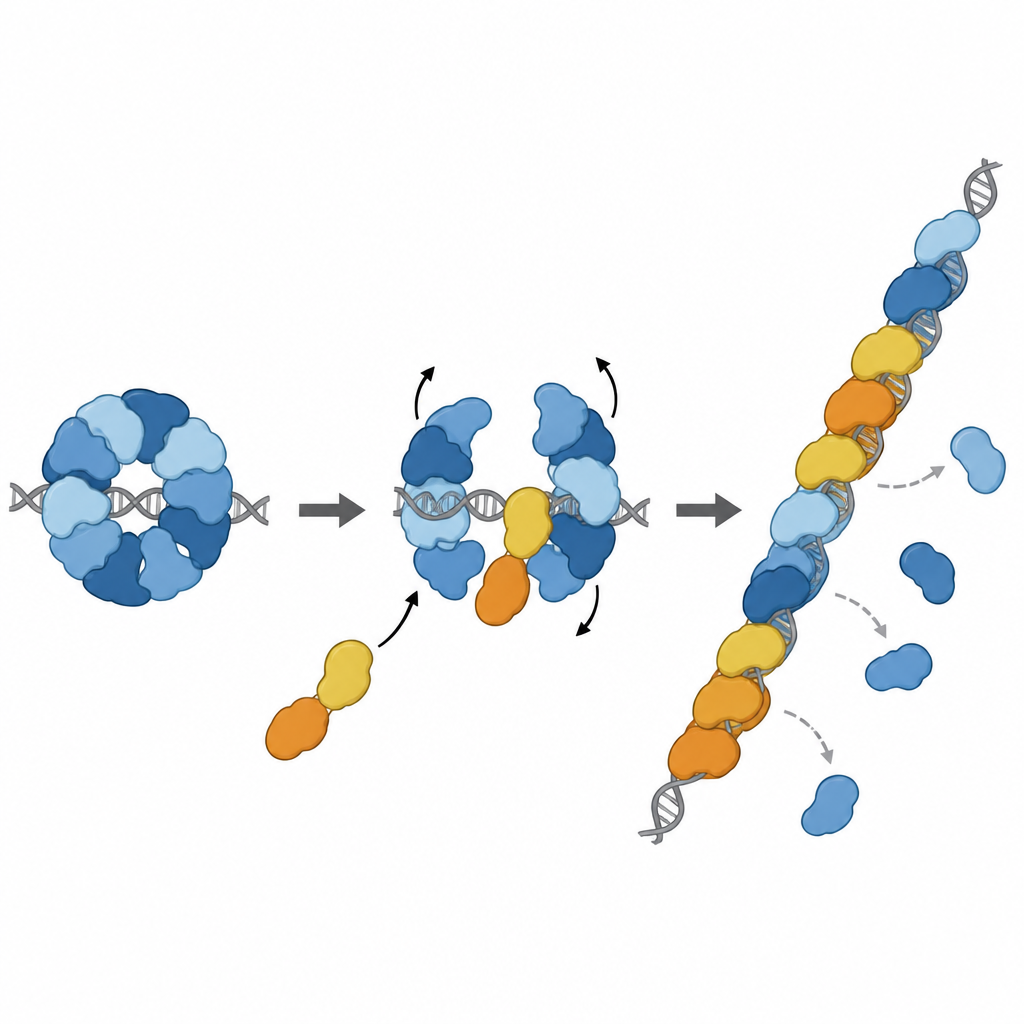
Присоединение к ремонтной линии в нужном месте
Дальнейшие структурные исследования показали комплекс RAD51–X3CDX2 непосредственно на конце филамента RAD51 на одноцепочечной ДНК. Здесь паралоги фиксируют пятимировой конец филамента и тонко меняют удержание соседних оснований ДНК, нарушая обычный повторяющийся триплетный узор, наблюдаемый в филаментах только из RAD51. Это говорит о том, что комплекс как инициирует рост филамента, так и стабилизирует растущий конец. Эксперименты в клетках подтвердили важность этих структурных особенностей: мутации, разрушающие ключевые точки контакта XRCC3 с RAD51, ослабляли ремонт ДНК посредством гомологичной рекомбинации и нарушали рестарт остановленных репликационных вилок — процессы, жизненно важные для поддержания стабильности генома.
Что это значит для здоровья и болезни
Вместе эти результаты выявляют двухступенчатую систему контроля для центрального фактора ремонта ДНК. Сначала комплекс RAD51–X3C удерживает RAD51 подальше от бесполезных двухцепочечных или РНК–ДНК участков, направляя его к разорванной одноцепочечной ДНК. Затем, когда условия благоприятны, приходит DX2 и перестраивает этот ансамбль в активный блок RAD51–X3CDX2, который инициирует и стабилизирует ремонтные филаменты. Поскольку наследственные изменения в нескольких из этих паралогов связаны с раком молочной железы, яичников и другими опухолями, понимание этого молекулярного «передачи» даёт более ясную картину того, как обычно сохраняется стабильность генома и как её нарушение может приводить к болезни.
Цитирование: Rawal, Y., Kwon, Y., Jia, L. et al. Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation. Nat Struct Mol Biol 33, 768–781 (2026). https://doi.org/10.1038/s41594-026-01796-6
Ключевые слова: Ремонт ДНК, RAD51, гомологичная рекомбинация, репликационный стресс, генетика рака