Clear Sky Science · de
Strukturelle Einblicke, wie der Austausch von RAD51-Paralogen die Bildung von RAD51-Filamenten reguliert
Unsere DNA vor alltäglichen Schäden schützen
Jede Zelle Ihres Körpers kopiert und repariert ständig ihre DNA, dennoch bleibt das genetische Programm über ein Leben hinweg bemerkenswert stabil. Versagt dieses Reparatursystem, können sich Mutationen ansammeln und zur Entstehung von Krebs beitragen. Dieser Artikel beleuchtet, wie eine Gruppe von Helferproteinen einen entscheidenden Schritt der DNA-Reparatur choreografiert und damit eine molekulare Sicherheitsvorkehrung offenbart, die effiziente Reparatur fördert und unnötige Umwege vermeidet.
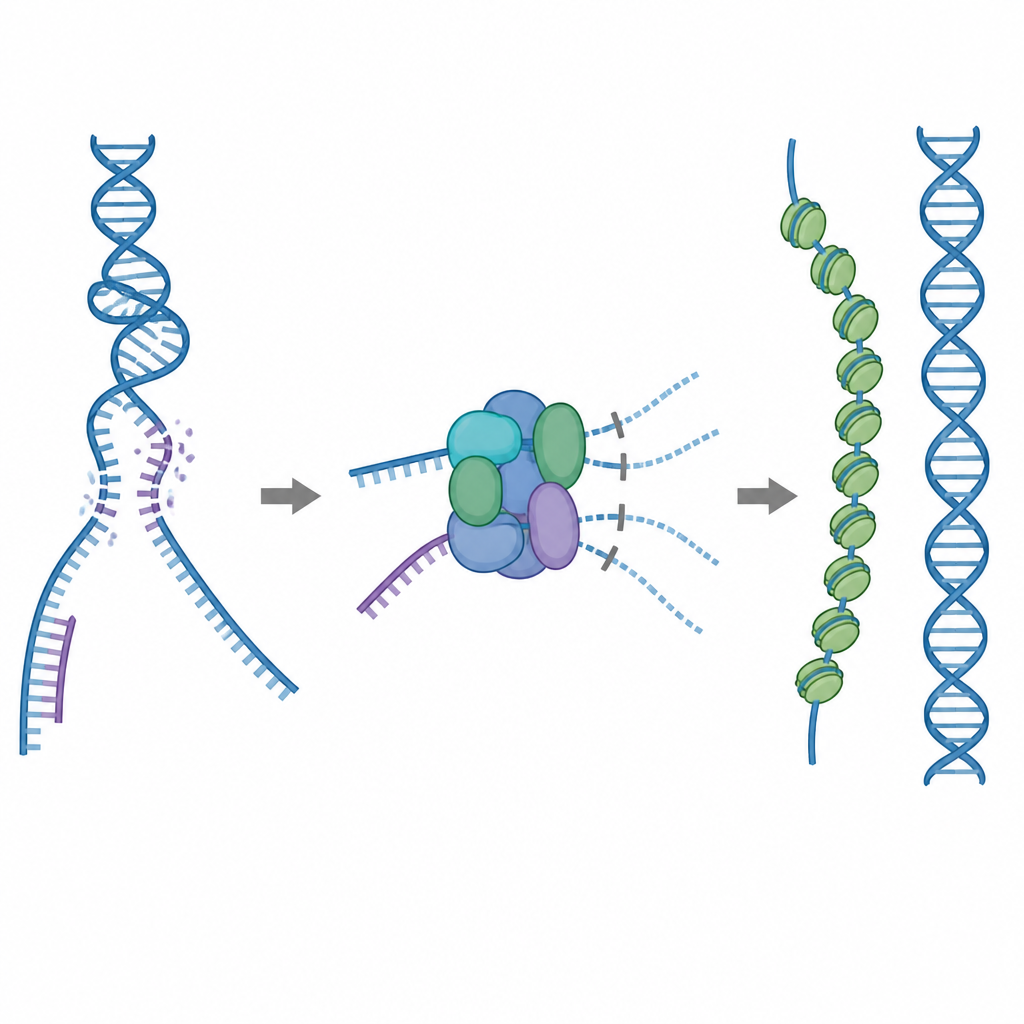
Ein Reparaturwerkzeug, das mit Vorsicht eingesetzt werden muss
Eines der wichtigsten Reparaturwerkzeuge der Zelle ist das Protein RAD51, das lange Filamente auf einzelsträngiger, gebrochener DNA bildet. Diese Filamente suchen nach einer übereinstimmenden Sequenz auf einer intakten DNA-Kopie und vermitteln den Strangtausch, damit der Bruch präzise repariert werden kann. RAD51 kann sich jedoch auch an falsche Ziele binden, etwa an normale doppelsträngige DNA oder an RNA–DNA-Hybride. Wenn zu viel RAD51 an solchen unproduktiven Stellen gebunden ist, bleiben echte Brüche möglicherweise unrepariert oder werden durch toxische Protein–DNA-Verwicklungen blockiert. Um das zu verhindern, nutzen Zellen fünf verwandte Helferproteine, die als RAD51-Paraloge bekannt sind, deren genaue Aufgaben jedoch lange unklar waren.
Eine eingebaute Bremse, die verschwendetes Binden verhindert
Mithilfe der Kryo-Elektronenmikroskopie visualisierten die Forscher, wie RAD51 mit zwei Paralogen, RAD51C und XRCC3, ein Komplex namens RAD51–X3C bildet. Sie fanden heraus, dass sich acht Proteinuntereinheiten zu einer ringförmigen Struktur zusammenfügen, angeordnet als zwei Tetramere. In diesem Zustand nimmt eine spezielle Schleife von XRCC3 physisch die Oberfläche von RAD51 ein, die normalerweise DNA greift, und benachbarte RAD51-Subunits sind aus der Ausrichtung verdreht. Biochemische Tests zeigten, dass dieser ‚autoinhibierte‘ Komplex einzelsträngige DNA noch recht gut bindet, seine Fähigkeit, an doppelsträngige DNA oder RNA–DNA-Hybride zu binden, jedoch stark reduziert ist. Effektiv wirkt der Komplex wie eine Bremse, die verhindert, dass RAD51 in unproduktive Bindungsstellen gelockt wird, während er gleichzeitig frei bleibt, die einzelsträngigen Regionen anzusprechen, die für die präzise Reparatur nötig sind.
Die Bremse in einen Beschleuniger verwandeln
Die Geschichte endet nicht beim Bremsen. Zwei weitere Paraloge, RAD51D und XRCC2, bilden ein Partnerkomplex namens DX2. Das Team zeigte, dass DX2 an RAD51–X3C andocken kann, indem es an einer exponierten Fläche von RAD51C ansetzt. Dieser ‚Paralogenaustausch‘ baut den achtteiligen Ring in eine Fünf-Teile-Anordnung um, RAD51–X3CDX2. In dieser neuen Form ist die DNA greifende Fläche von RAD51 freigelegt und korrekt ausgerichtet, wodurch eine durchgehende, filamentfähige Oberfläche entsteht. Der umgestaltete Komplex bindet einzelsträngige DNA stärker, unterstützt den Aufbau von RAD51-Filamenten auf langen Strängen und kann – wichtig – RAD51 auch dann laden, wenn diese Stränge zunächst von einem anderen Schutzprotein namens RPA bedeckt sind. In in-vitro-Strangtausch-Experimenten stellte der pentamerische Komplex die Aktivität von RAD51 unter Bedingungen wieder her, unter denen RAD51 allein stark gehemmt war.
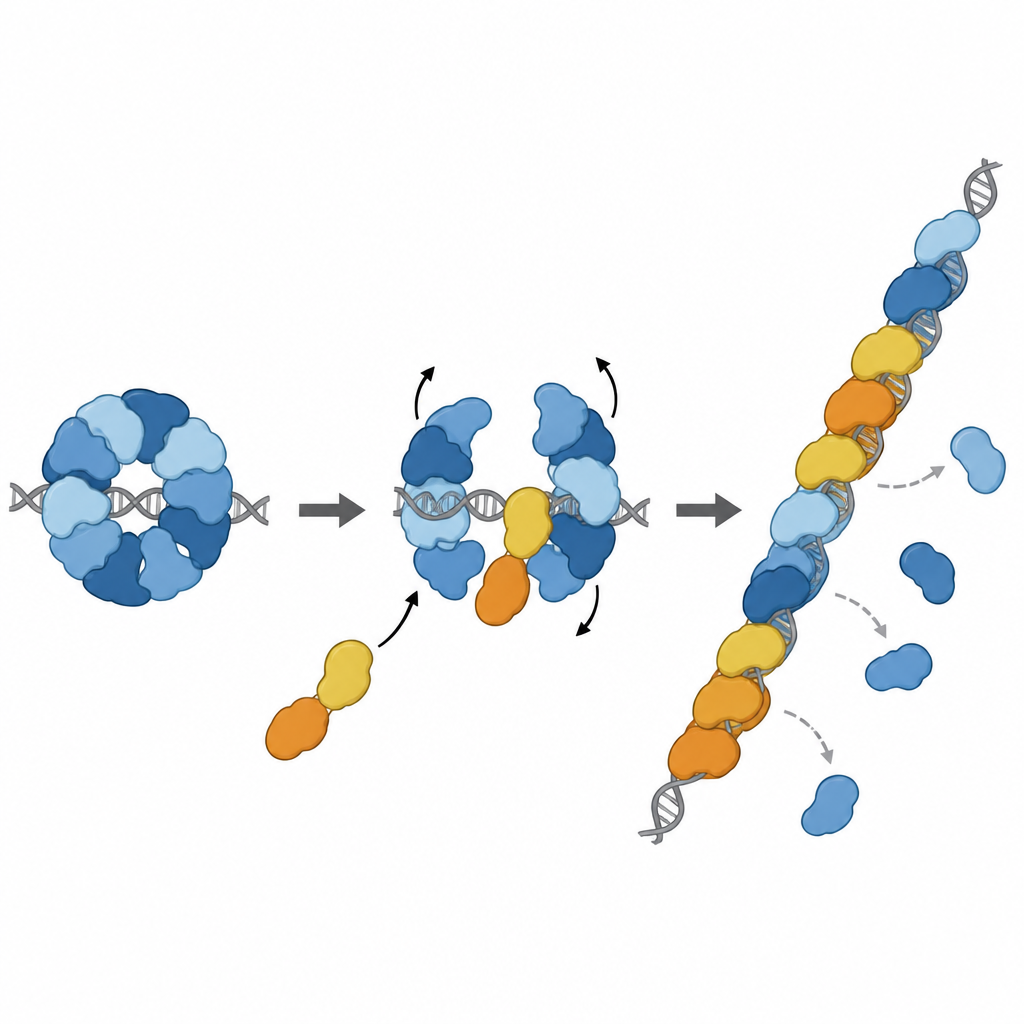
Genau an der richtigen Stelle in die Reparaturkette einsteigen
Weitere strukturelle Untersuchungen zeigten den RAD51–X3CDX2-Komplex tatsächlich am Ende eines RAD51-Filaments auf einzelsträngiger DNA. Hier kappten die Paraloge das 5'-Ende des Filaments und veränderten subtil, wie umliegende DNA-Basen gehalten werden, wodurch das sonst bei RAD51-allein-Filamenten sichtbare sich wiederholende Tripletmuster gebrochen wurde. Das legt nahe, dass der Komplex sowohl das Ansprossen des Filamentwachstums initiiert als auch das wachsende Ende stabilisiert. Zellbasierte Experimente unterstützten die Bedeutung dieser strukturellen Merkmale: Mutationen, die die Schlüsselkontaktpunkte von XRCC3 mit RAD51 stören, schwächten die homologe Rekombinationsreparatur und beeinträchtigten das Wiederanlaufen gestoppter DNA-Replikationsgabeln – Prozesse, die für die Aufrechterhaltung der Genomstabilität entscheidend sind.
Bedeutung für Gesundheit und Krankheit
Zusammen zeigen diese Ergebnisse ein zweistufiges Kontrollsystem für einen zentralen DNA-Reparaturfaktor. Zuerst hält der RAD51–X3C-Komplex RAD51 von unproduktiven doppelsträngigen oder RNA–DNA-Regionen fern und lenkt es zu gebrochener, einzelsträngiger DNA. Wenn die Bedingungen stimmen, trifft dann DX2 ein und remodeliert diesen Komplex zu einer aktiven RAD51–X3CDX2-Einheit, die Reparaturfilamente ansiedelt und stabilisiert. Da vererbte Veränderungen in mehreren dieser Paraloge mit Brust-, Eierstock- und anderen Krebsarten verknüpft sind, liefert das Verständnis dieses molekularen Übergangs ein klareres Bild davon, wie die Genomstabilität normalerweise bewahrt wird und wie ihr Zusammenbruch bei Erkrankungen ablaufen kann.
Zitation: Rawal, Y., Kwon, Y., Jia, L. et al. Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation. Nat Struct Mol Biol 33, 768–781 (2026). https://doi.org/10.1038/s41594-026-01796-6
Schlüsselwörter: DNA-Reparatur, RAD51, homologe Rekombination, Replikationsstress, Krebsgenetik