Clear Sky Science · sv
Strukturell insikt i hur utbyte av RAD51‑paraloger reglerar RAD51‑filamentbildning
Att hålla vårt DNA säkert från vardaglig skada
Varje cell i din kropp kopierar och reparerar ständigt sitt DNA, ändå förblir den genetiska texten anmärkningsvärt stabil över en livstid. När detta reparationssystem sviktar kan mutationer ackumuleras och bidra till cancer. Den här artikeln undersöker hur en uppsättning hjälpproteiner koreograferar ett avgörande steg i DNA‑reparationen och avslöjar en molekylär säkerhetsmekanism som håller reparationen effektiv och undviker slöseri med felaktiga avvägar.
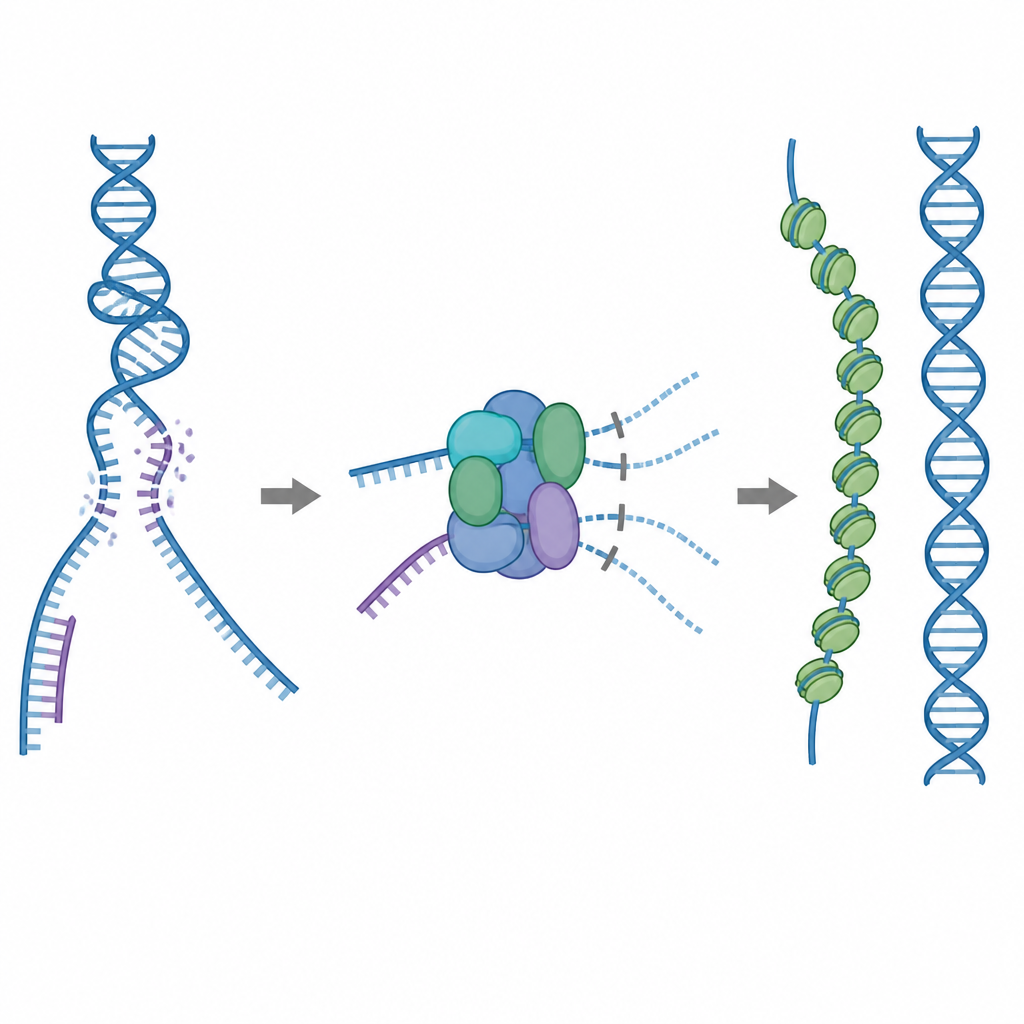
Ett reparationsverktyg som måste användas varsamt
Ett av cellens huvudverktyg för reparation är ett protein som kallas RAD51, vilket bildar långa filament på enkelsträngat brutet DNA. Dessa filament söker efter en matchande sekvens på en intakt DNA‑kopia och hjälper till att byta strängar så att brottet kan lagas korrekt. Men RAD51 kan också fästa vid fel mål, såsom normalt dubbelsträngt DNA eller strukturer som innehåller både RNA och DNA. Om för mycket RAD51 binds upp i dessa oproduktiva platser kan de verkliga brotten lämnas oreparerade eller belamrade med toxisk protein–DNA‑trassel. För att förhindra detta använder celler fem närbesläktade hjälpproteiner, kända som RAD51‑paraloger, vars exakta roller hittills varit oklara.
En inbyggd broms som blockerar slösaktig bindning
Genom kryoelektronmikroskopi visualiserade forskarna hur RAD51 slår ihop sig med två paraloger, RAD51C och XRCC3, för att bilda ett komplex kallat RAD51–X3C. De fann att åtta proteinenheter går ihop i en ringliknande struktur, arrangerad som två tetramerer. I detta tillstånd upptar en särskild slingform från XRCC3 fysiskt den yta på RAD51 som normalt greppar DNA, och närliggande RAD51‑underenheter vrids ur linje. Biokemiska tester visade att detta ”autoinhiberade” komplex fortfarande binder enkelsträngat DNA hyfsat bra, men dess förmåga att fästa vid dubbelsträngat DNA eller RNA–DNA‑hybrider är kraftigt reducerad. I praktiken fungerar komplexet som en broms som hindrar RAD51 från att sugas upp av ickeproduktiva bindningsställen samtidigt som det lämnas fritt att engagera de enkelsträngar som behövs för korrekt reparation.
Att förvandla bromsen till en förstärkare
Berättelsen slutar inte vid bromsning. Två andra paraloger, RAD51D och XRCC2, bildar ett partnerkomplex kallat DX2. Teamet visade att DX2 kan ansluta till RAD51–X3C genom att docka på en exponerad yta av RAD51C. Detta ”paralogutbyte” omformar den åtta‑delade ringen till en femdelad uppställning, RAD51–X3CDX2. I denna nya form blottas RAD51:s DNA‑greppande yta och linjeras upp korrekt, vilket skapar en kontinuerlig yta redo för filamentbildning. Det ombyggda komplexet binder enkelsträngat DNA tätare, hjälper RAD51 att montera filament på långa strängar och, vilket är viktigt, kan ladda RAD51 även när dessa strängar initialt är täckta av ett annat skyddande protein som kallas RPA. I laboratoriebaserade strängbyte‑reaktioner återställde det pentameriska komplexet RAD51:s aktivitet under förhållanden där enbart RAD51 var starkt hämmat.
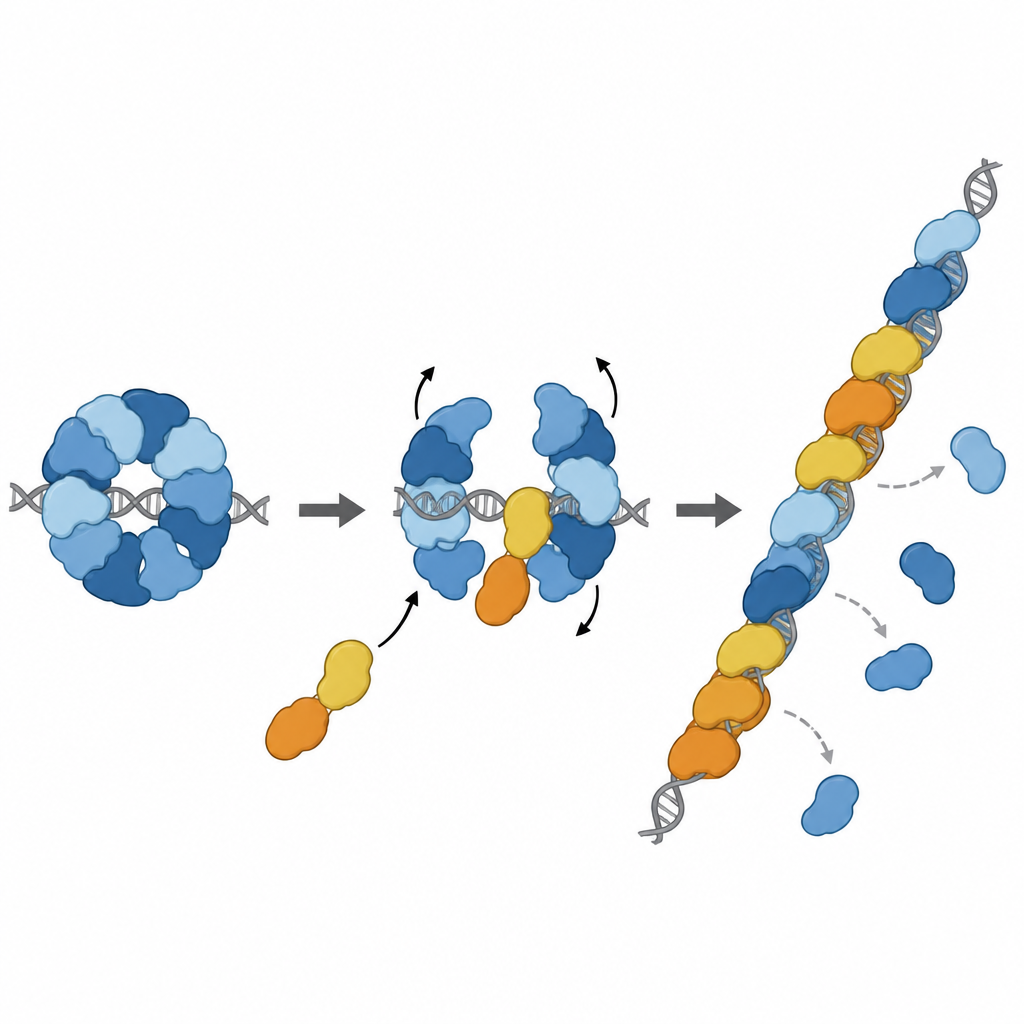
Att gå med i reparationskedjan på precis rätt plats
Ytterligare strukturarbete fångade RAD51–X3CDX2‑komplexet sittande i änden av ett RAD51‑filament på enkelsträngat DNA. Här kapslar paralogerna filamentets 5′‑ände och ändrar subtilt hur närliggande DNA‑baser hålls, vilket bryter det vanliga upprepade tripletmönstret som ses i filament som endast innehåller RAD51. Detta tyder på att komplexet både initierar filamenttillväxt och stabiliserar den växande änden. Cellbaserade experiment stödde vikten av dessa strukturella drag: mutationer som stör XRCC3:s centrala kontaktpunkter med RAD51 försvagade DNA‑reparation via homolog rekombination och försämrade återstarten av fastnade DNA‑replikationsgafflar, processer som är avgörande för att bevara genomets stabilitet.
Vad detta betyder för hälsa och sjukdom
Tillsammans avslöjar dessa fynd ett tvåstegskontrollsystem för en central DNA‑reparationsfaktor. Först håller RAD51–X3C‑komplexet RAD51 borta från oönskade dubbelsträngade eller RNA–DNA‑regioner och styr det mot brutet, enkelsträngat DNA. Sedan, när förhållandena är riktiga, anländer DX2 för att omforma denna sammansättning till en aktiv RAD51–X3CDX2‑enhet som initierar och stabiliserar reparationsfilament. Eftersom ärftliga förändringar i flera av dessa paraloger kopplats till bröst‑, äggstocks‑ och andra cancerformer, ger förståelsen av denna molekylära överlämning en tydligare bild av hur genomstabilitet normalt bevaras och hur dess sammanbrott kan ske vid sjukdom.
Citering: Rawal, Y., Kwon, Y., Jia, L. et al. Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation. Nat Struct Mol Biol 33, 768–781 (2026). https://doi.org/10.1038/s41594-026-01796-6
Nyckelord: DNA‑reparation, RAD51, homolog rekombination, replikationsstress, cancergenetik