Clear Sky Science · es
Perspectiva estructural sobre cómo el intercambio de paralogos de RAD51 regula la formación del filamento de RAD51
Proteger nuestro ADN frente al daño cotidiano
Cada célula de tu cuerpo copia y repara constantemente su ADN, y, sin embargo, el guion genético permanece notablemente estable a lo largo de la vida. Cuando este sistema de reparación falla, las mutaciones pueden acumularse y contribuir al cáncer. Este artículo explora cómo un conjunto de proteínas auxiliares coreografía un paso crucial de la reparación del ADN, revelando un mecanismo molecular que mantiene la reparación eficiente y evita desvíos inútiles.
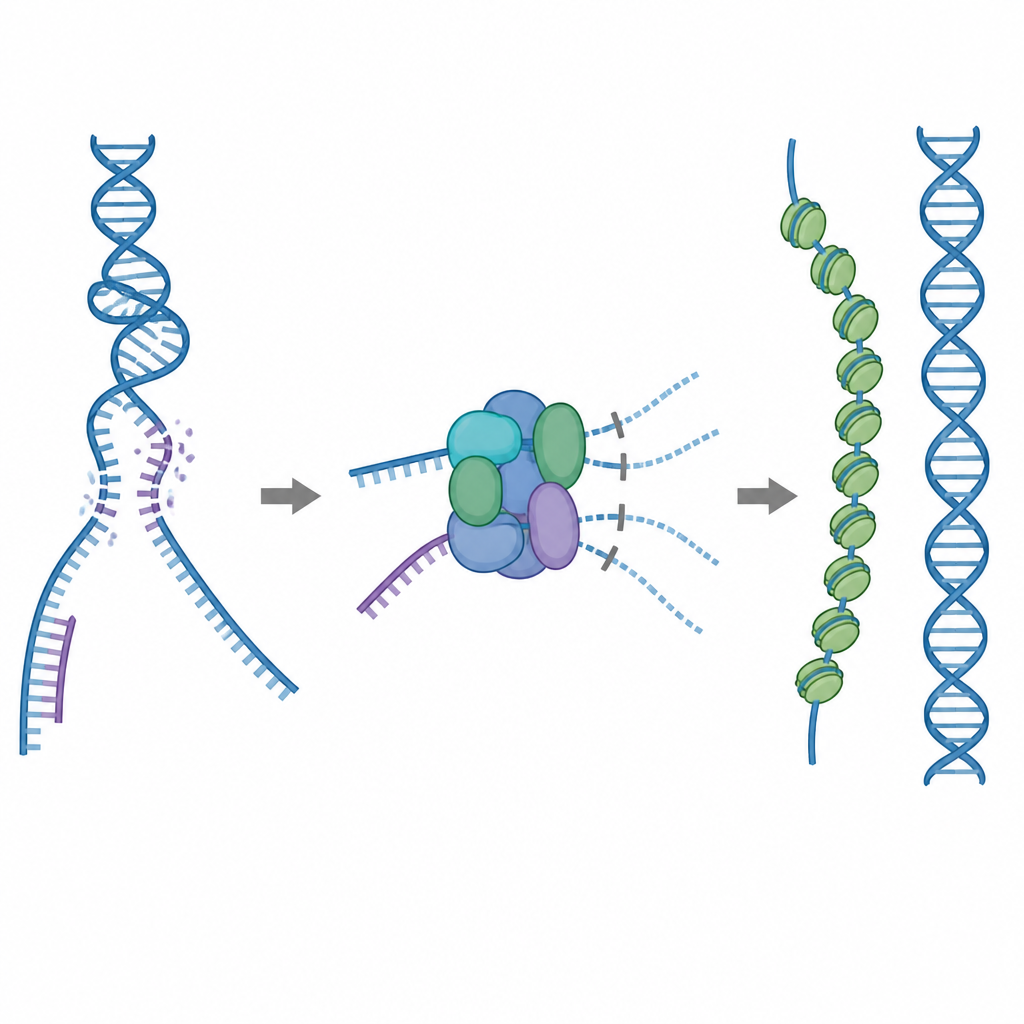
Una herramienta de reparación que debe usarse con cuidado
Una de las principales herramientas de reparación de la célula es una proteína llamada RAD51, que forma filamentos largos sobre hebras simples de ADN roto. Estos filamentos buscan una secuencia coincidente en una copia intacta del ADN y facilitan el intercambio de hebras para que la rotura pueda repararse con precisión. Pero RAD51 también puede adherirse a blancos equivocados, como ADN de doble hebra normal o estructuras que contienen híbridos ARN–ADN. Si demasiado RAD51 queda atrapado en esos sitios no útiles, las roturas reales pueden quedarse sin reparar o congestionadas con enredos tóxicos proteína–ADN. Para evitar esto, las células usan cinco proteínas auxiliares relacionadas, conocidas como paralogos de RAD51, cuyos roles detallados no habían quedado claros.
Un freno incorporado que bloquea uniones improductivas
Mediante microscopía crioelectrónica, los investigadores visualizaron cómo RAD51 se asocia con dos paralogos, RAD51C y XRCC3, para formar un complejo llamado RAD51–X3C. Encontraron que ocho unidades proteicas se unen en una estructura anular, dispuestas como dos tetrámeros. En este estado, un lazo especial de XRCC3 ocupa físicamente la superficie de RAD51 que normalmente se engancha al ADN, y las subunidades vecinas de RAD51 quedan torcidas y desalineadas. Pruebas bioquímicas mostraron que este complejo “autoinhibido” aún se une razonablemente bien al ADN de hebra sencilla, pero su capacidad para adherirse al ADN de doble hebra o a híbridos ARN–ADN queda fuertemente reducida. En efecto, el complejo actúa como un freno que evita que RAD51 se absorba en sitios de unión no productivos, a la vez que lo deja libre para actuar sobre las hebras simples necesarias para una reparación precisa.
Convertir el freno en un impulsor
La historia no termina con el freno. Otros dos paralogos, RAD51D y XRCC2, forman un complejo asociado llamado DX2. El equipo mostró que DX2 puede unirse a RAD51–X3C acoplándose a una superficie expuesta de RAD51C. Este “intercambio de paralogos” remodela el anillo de ocho partes hacia una disposición de cinco componentes, RAD51–X3CDX2. En esta nueva forma, la cara de RAD51 que agarra el ADN queda descubierta y alineada correctamente, generando una superficie continua apta para formar filamentos. El complejo remodelado se une con mayor afinidad al ADN de hebra sencilla, ayuda a RAD51 a ensamblar filamentos sobre largas hebras y, de forma importante, puede cargar RAD51 aun cuando esas hebras estén inicialmente recubiertas por otra proteína protectora llamada RPA. En reacciones de intercambio de hebras en el laboratorio, el complejo pentamérico restauró la actividad de RAD51 en condiciones donde RAD51 solo estaba fuertemente inhibido.
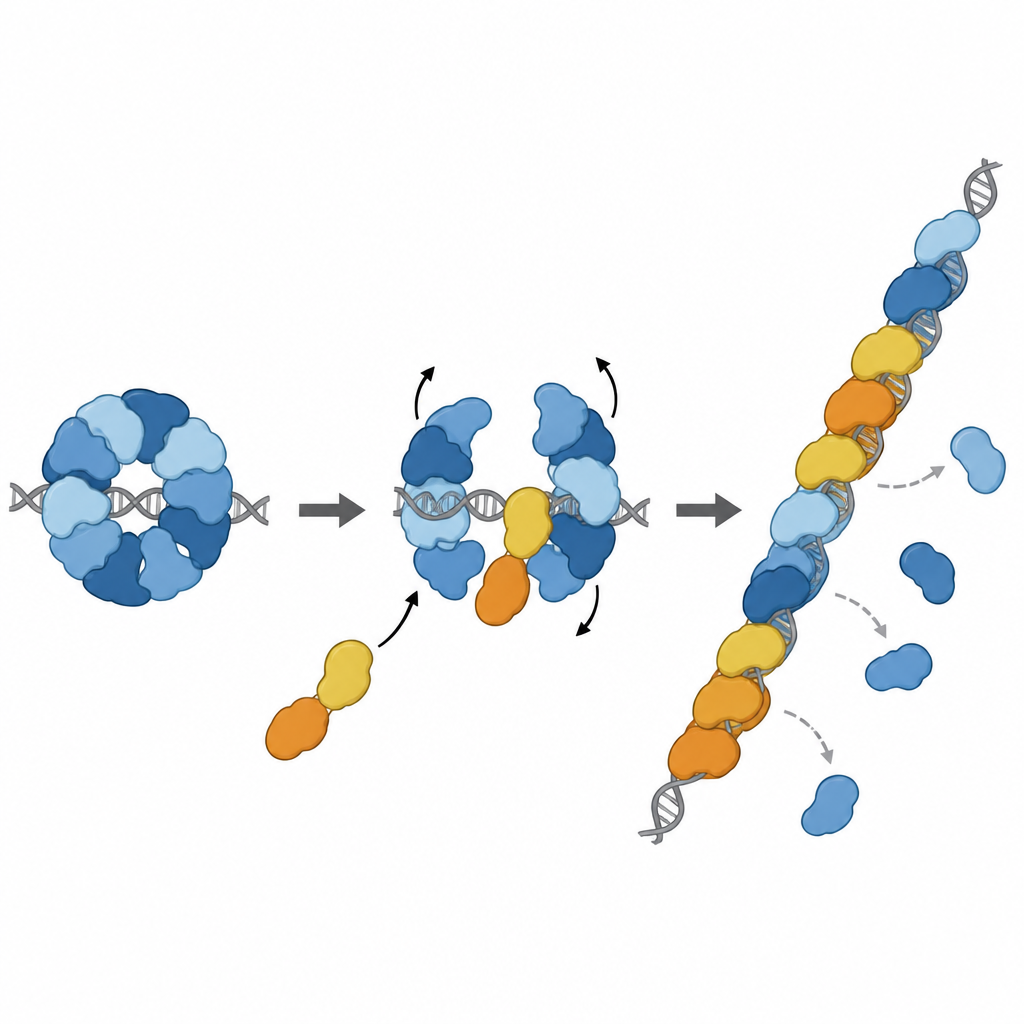
Incorporarse a la reparación en el lugar adecuado
Trabajos estructurales adicionales capturaron al complejo RAD51–X3CDX2 realmente situado en el extremo de un filamento de RAD51 sobre ADN de hebra sencilla. Allí, los paralogos tapan el extremo 5' del filamento y alteran sutilmente cómo se sujetan las bases de ADN cercanas, rompiendo el patrón repetitivo de tripletes que se observa en filamentos formados solo por RAD51. Esto sugiere que el complejo tanto inicia el crecimiento del filamento como estabiliza el extremo en expansión. Experimentos en células respaldaron la importancia de estas características estructurales: mutaciones que interrumpen los puntos de contacto clave de XRCC3 con RAD51 debilitan la reparación por recombinación homóloga y perjudican el reinicio de las horquillas de replicación detenidas, procesos vitales para mantener la estabilidad del genoma.
Qué significa esto para la salud y la enfermedad
En conjunto, estos hallazgos revelan un sistema de control en dos pasos para un factor central de la reparación del ADN. Primero, el complejo RAD51–X3C mantiene a RAD51 alejado de regiones no útiles de doble hebra o híbridos ARN–ADN, dirigiéndolo hacia el ADN roto de hebra sencilla. Luego, cuando las condiciones son adecuadas, DX2 llega para remodelar este ensamblaje en una unidad activa RAD51–X3CDX2 que inicia y estabiliza los filamentos de reparación. Dado que cambios heredados en varios de estos paralogos están vinculados al cáncer de mama, ovario y otros, comprender este relevo molecular ofrece una imagen más clara de cómo se preserva normalmente la estabilidad del genoma y de cómo su fallo puede contribuir a la enfermedad.
Cita: Rawal, Y., Kwon, Y., Jia, L. et al. Structural insight into how RAD51 paralog exchange regulates RAD51 filament formation. Nat Struct Mol Biol 33, 768–781 (2026). https://doi.org/10.1038/s41594-026-01796-6
Palabras clave: Reparación del ADN, RAD51, recombinación homóloga, estrés de replicación, genética del cáncer