Clear Sky Science · zh
β-内酰胺酶 TEM-1 α-螺旋中双突变表位互作的起源与广度
为什么蛋白质的微小变化会影响抗生素耐药性
耐药细菌的威胁日益加剧,许多细菌依赖能够在药物发挥作用前将其破坏的酶。其中一种叫做 TEM-1 β-内酰胺酶的酶,帮助常见肠道细菌抵抗像阿莫西林这样的常用抗生素。本研究聚焦于该酶中的一个极小片段——由11个氨基酸构成的 α-螺旋——提出一个重要问题:成对的突变如何相互作用以使耐药性更容易或更困难地演化?我们能否从基本物理原理和进化数据中预测这些效应?
当一个突变依赖于另一个时
这里研究的核心概念是“表位互作”(epistasis),意思是一个突变的效应取决于其它突变是否存在。突变并非独立作用,它们可以以出人意料的方式相互促进或抵消。作者构建了一个包含超过14,000种变体的大型文库,每种变体在同一 α-螺旋内携带一个或两个突变,该螺旋虽不在活性位点但有助于维持蛋白构象。然后他们用两种方式测量每个变体对细菌抵抗阿莫西林的保护作用:通过竞争生长追踪细菌生长(直接的适应度测量)和测定抑制生长的最低抗生素浓度(MIC,临床标准的耐药性测量)。这两种配对测量使他们能够量化突变效应何时是依赖背景的,而不是简单相加的。
许多突变会破坏酶活性,但大多数相互作用可预测
第一个显著发现是这个小螺旋非常脆弱。在测试条件下,该区域近一半的单个突变基本破坏了酶的功能,而近五分之四的双突变体是无功能的。某些替换,例如引入脯氨酸,几乎总是致死,因为它会破坏螺旋的结构。当研究团队将双突变的表现与单个突变效应相加的预期比较时,发现普遍存在表位互作:剔除真正失活的变体后,大约90%的突变表现出明显的背景依赖性。然而,这些表位互作大多呈现简单模式——突变在已经被削弱的背景上往往看起来更糟,这是一种“全局”负性互作。
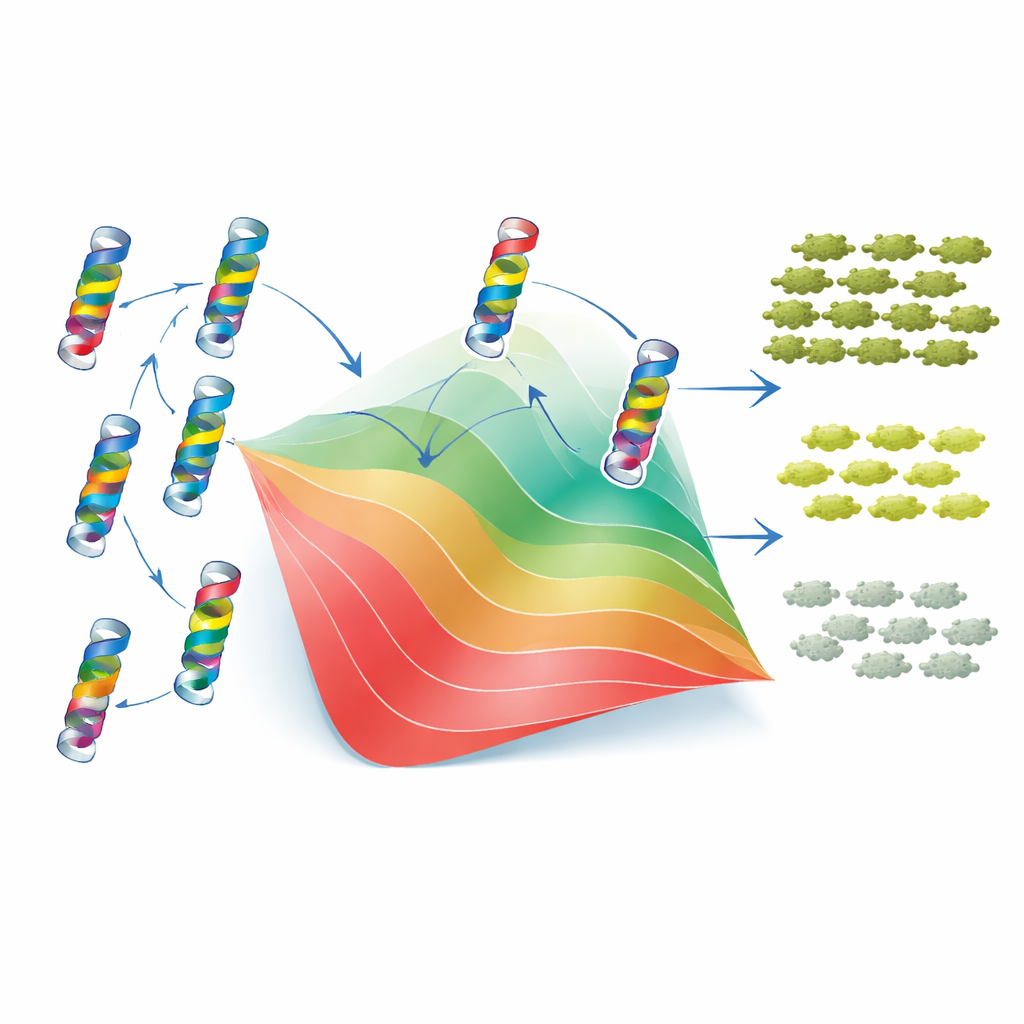
一个简单的折叠模型解释了大部分复杂性
为理解这些模式为何出现,作者采用了经典的两态蛋白折叠模型。在该视角下,TEM-1 要么折叠并有功能,要么未折叠且无用,每个突变通过提高或降低蛋白稳定性来改变两种状态的平衡。关键假设是,单个突变引起的稳定性变化可以相加,尽管最终对适应度的影响是非线性的:一旦蛋白足够不稳定,微小的额外变化会带来更大的后果。通过将该框架拟合到实验数据,研究者可以为每个突变分配一个有效的“稳定性代价”或“稳定性增益”。仅凭这些值和一个基线稳定性参数,模型就能准确再现数千种单、双突变体的适应度和 MIC,以及表位互作的总体分布和偏向。换言之,突变相互作用的许多表面复杂性源于一个简单的物理约束:蛋白必须保持折叠状态。
局部接触与长期进化留下更微妙的印记
但故事并非完全由全局效应主导。对于在三维结构中彼此非常接近的残基对,两态模型的解释效果较差。对于这些邻近残基,数据揭示了“特异性”表位互作——某些特定组合的表现比仅靠稳定性预期的更好或更差,有时它们会在电荷或形状上相互补偿。为了检验进化是否保留了这些局部互作的痕迹,作者使用统计性的“Potts”模型分析了成千上万条相关的 β-内酰胺酶序列,该模型刻画了哪些残基在物种间倾向共同变异。这些基于序列的耦合与实验发现有两方面的一致性:它们与推断出的稳定性效应相关联,并突出了许多简单折叠模型失效的残基对。当对 Potts 模型的输出进行适当重标度并通过相同的折叠框架处理时,它们甚至能部分预测新突变对的表位互作符号(正或负)。
这对预测耐药性的意义
对非专业读者而言,关键结论是:即便在单个蛋白的一个微小结构元件内,突变之间的相互作用也很常见,但并非随机。大多数可以被理解为突变如何集体将酶推向或远离稳定折叠状态的结果,而少数反映直接的局部接触以微调性能。两类效应都在 β-内酰胺酶的长期进化中留下了可检测的痕迹,表明突变相互作用的规则足够稳定,可以被学习和推广。这意味着,通过将深度突变扫描与蛋白折叠模型和进化序列数据相结合,最终可能预测耐药酶如何演化——并设计出更难被其突破的药物或疗法。
引用: Birgy, A., Roussel, C., Kemble, H. et al. Origins and breadth of pairwise epistasis in an α-helix of β-lactamase TEM-1. Nat Commun 17, 4083 (2026). https://doi.org/10.1038/s41467-026-70627-5
关键词: 表位互作, 蛋白质稳定性, β-内酰胺酶, 抗生素耐药性, 深度突变扫描