Clear Sky Science · zh
光能驱动的鞭毛微生物美洲眼虫的 PSI–LHCI–LHCII 架构与能量传递通路
微小游动生物如何将阳光变为动力
单细胞藻类美洲眼虫(Euglena gracilis)是栖息于池塘与水坑的微小游动生物。像植物一样,它利用阳光维持生命,但其光合作用途径走出了一条不同寻常的进化道路。本研究以原子级别揭示了其光合作用机器,展示了美洲眼虫在数百万年前通过获得另一藻类的叶绿体后,如何重构其光捕获系统以实现高效且灵活的能量捕获。
来自“借来”的叶绿体的特殊光学引擎
光合作用的核心是光系统 I(Photosystem I),一种将光能转化为推动糖类合成所需电子流的蛋白-色素复合体。在大多数植物和绿藻中,这一核心单元周围环绕着新月形的天线蛋白负责收集光。美洲眼虫则不同:它的叶绿体源自吞噬了一个已经会进行光合作用的绿藻,属于生物学上所称的次级质体。这样非同寻常的来源使美洲眼虫保留了独特的色素和蛋白成分,但其光系统 I 超复合体的精确布局直到现在尚不清楚。作者借助高分辨率冷冻电镜解析出该组装体的三维结构,分辨率达2.06埃,揭示了美洲眼虫如何重建它的光学引擎。
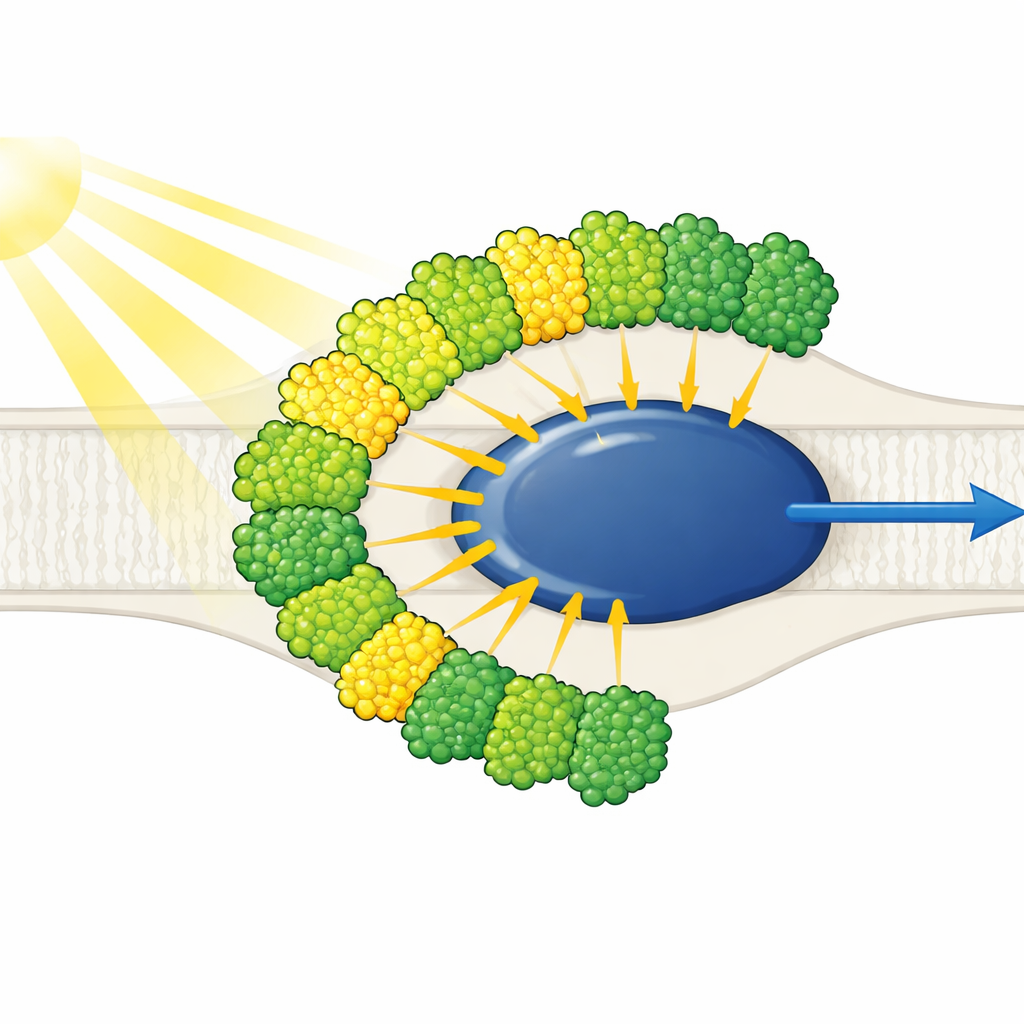
所有光天线都集中在一侧
结构显示,简化的光系统 I 核心被十个光捕获天线包围——九个为一种类型(LHCI),另有一个通常与光系统 II 相关的单元(LHCII)。显著的是,所有十个天线都位于核心的同一侧,排列为两个同心带,而不是像植物那样在对侧形成新月形。每个外层天线与内层相邻单元面对面配对,形成五个堆叠二聚体,这些二聚体进一步组装成两个四聚体。这些天线由多肽前体切割成单个单位构成,特定的短序列基序像分子魔术贴一样将二聚体粘合在一起。一个称为 LHC-3 的内层天线成为中心枢纽,连接二聚体并帮助将整个天线带固定到核心上。
替代丢失部件的新蛋白环
与常见的植物系统相比,美洲眼虫的光系统 I 核心缺失了若干通常用于定位天线并在光照变化时实现动态重组的小锚定蛋白。作为补偿,其剩余的核心亚基之一 PsaD 带有约40个氨基酸的额外环段,这段插入仅见于美洲眼虫。该灵活插入伸入缺失蛋白原本所在的空间,并与 LHC-3 及其叶绿素形成直接的稳定接触。作者提出,这一独特环段可能与丢失亚基的演化同时发生,取代它们固定天线与核心的作用,并决定入射光能进入机器的路径与位置。
定制色素与能量高速公路
除了蛋白质的整体结构外,叶绿素分子的排列决定了光能的流动方式。美洲眼虫复合体携带200多个色素,包括叶绿素 a 与 b 以及与陆生植物不同的特殊类胡萝卜素。按已知位点类比标记为 a1020 和 a1021 的两枚叶绿素是美洲眼虫所特有,仅出现在内层天线上。它们位于三枚天线交汇处,形成连接相邻单元的色素簇。这些簇连同紧密配对的“红色”叶绿素(吸收稍低能量光)创造了从外层带到内层带再进入核心的多个能量传递通路。基于色素中心间距离的计算表明,外层天线收集的能量优先被引导到少数门户天线,尤其是 LHC-3 及其对应的 LHC-9,然后再交给发生电子转移的反应中心。
用于适应变化光照的可动连接
并非所有天线都与核心牢固连接。冷冻电镜数据显示,独立的 LHCII 单元和一个外层 LHCI 仅呈部分占据,提示它们比其他天线更容易装配或脱落。这种可变性暗示了一种调节机制:通过增加或移除这些天线,美洲眼虫可能调整到达光系统 I 的光量,并改变与光系统 II 共享能量的平衡,类似植物所知的状态转换,但采用了不同的结构解决方案。结合其独特色素和不对称的天线排列,美洲眼虫的设计似乎在稳健与可调之间取得了优化。
对阳光驱动生命的意义
对非专业读者而言,核心信息是:进化有许多方法解决将阳光转化为可用能量的问题。美洲眼虫在以“二手”方式获得叶绿体后,并未简单复制标准的植物蓝图。相反,它简化了核心、将所有天线重定位到一侧、发明了新的蛋白环以锚定这些天线,并部署了独特的叶绿素簇与色素以高效引导能量进入反应中心。这一谱系特异的解决方案表明次级质体能够在重塑光合作用装置的同时保留——甚至增强——收获光能以维持生命的基本功能。
引用: Huang, G., Dong, S., Li, L. et al. The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis. Nat Commun 17, 3429 (2026). https://doi.org/10.1038/s41467-026-70183-y
关键词: 美洲眼虫, 光系统 I, 光捕获复合体, 激发能量传递, 冷冻电镜