Clear Sky Science · en
The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis
How a Tiny Swimmer Turns Sunlight into Power
The single-celled alga Euglena gracilis is a microscopic swimmer that thrives in ponds and puddles. Like plants, it uses sunlight to power life, but it does so with a photosynthetic machine that has taken an unusual evolutionary path. This study reveals that machine in atomic detail, showing how Euglena has rewired its light-collecting system to capture energy efficiently and flexibly after acquiring its chloroplast from another alga millions of years ago.
A Special Light Engine in a Borrowed Chloroplast
At the heart of photosynthesis lies Photosystem I, a protein–pigment complex that turns light energy into the flow of electrons needed to build sugars. In most plants and green algae, this core unit is surrounded by a crescent of antenna proteins that collect light. Euglena gracilis is different: its chloroplast originated by engulfing an already photosynthetic green alga, giving it what biologists call a secondary plastid. That unusual origin left Euglena with distinctive pigments and protein components, and until now, the exact layout of its Photosystem I supercomplex was unknown. Using high-resolution cryo-electron microscopy, the authors solved the three-dimensional structure of this assembly down to 2.06 angstroms, revealing how Euglena has rebuilt its light engine.
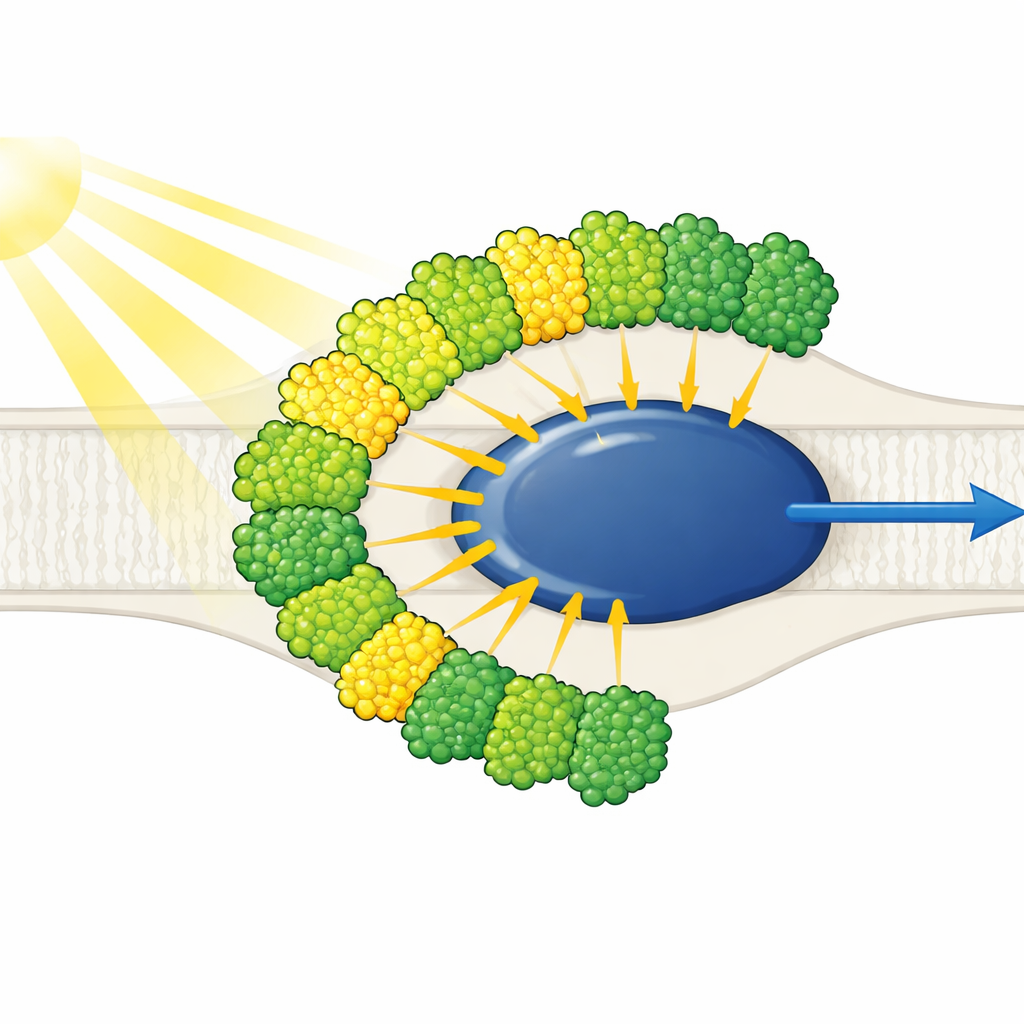
Light Antennas All on One Side
The structure shows a stripped-down Photosystem I core surrounded by ten light-harvesting antennas—nine of one type (LHCI) and one usually associated with Photosystem II (LHCII). Strikingly, all ten antennas sit on just one side of the core, in two concentric belts, rather than forming the typical plant-like crescent on the opposite side. Each outer antenna pairs face-to-face with an inner neighbor, creating five stacked dimers that in turn assemble into two tetramers. These antennas are built from polyproteins that are cut into individual units, and specific short sequence motifs act like molecular Velcro holding the dimers together. One inner antenna, called LHC-3, emerges as a central hub, linking dimers and helping to tie the entire belt to the core.
A New Protein Loop that Replaces Lost Parts
Compared with familiar plant systems, Euglena’s Photosystem I core is missing several small anchor proteins that usually help position antennas and enable dynamic reorganization under changing light. Instead, one of its remaining core subunits, PsaD, carries an extra loop of about 40 amino acids found only in Euglena. This flexible insertion reaches into the space where the missing proteins would normally sit and forms direct stabilizing contacts with LHC-3 and its chlorophylls. The authors propose that this unique loop evolved alongside the loss of the absent subunits, taking over their role in fastening antennas to the core and shaping where and how incoming light energy enters the machine.
Custom Pigments and Energy Highways
Beyond the architecture of proteins, the way chlorophyll molecules are positioned determines how light energy flows. The Euglena complex carries over 200 pigments, including chlorophylls a and b and special carotenoids that differ from those in land plants. Two chlorophylls, labeled a1020 and a1021 by analogy to known sites, are unique to Euglena and appear only in inner antennas. They sit at junctions where three antennas meet, forming pigment clusters that physically bridge neighboring units. These clusters, together with closely spaced “red” chlorophyll pairs that absorb slightly lower-energy light, create multiple routes for excitation to travel from the outer belt to the inner belt and then into the core. Calculations based on distances between pigment centers show that energy collected in outer antennas is funneled preferentially into a few gateway antennas, especially LHC-3 and its counterpart LHC-9, before being handed off to the reaction center where electron transfer occurs.
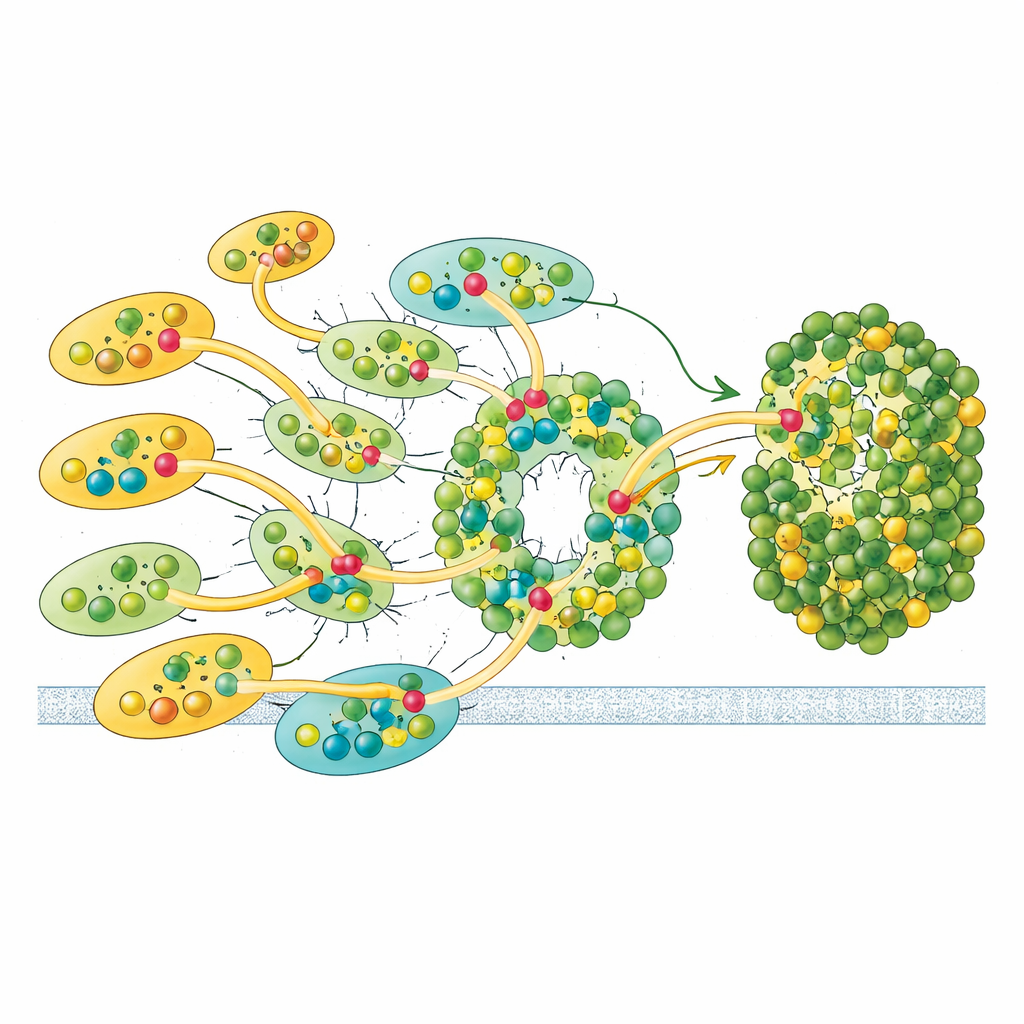
Flexible Links for Adapting to Changing Light
Not all antennas are bolted on equally tightly. The lone LHCII unit and one outer LHCI show only partial occupancy in the cryo-EM data, suggesting that they can attach and detach more readily than the others. This lability hints at a regulatory feature: by adding or removing these antennas, Euglena may tune how much light reaches Photosystem I and adjust the balance of energy shared with Photosystem II, echoing state transitions known from plants but using a different structural solution. Together with its unique pigments and asymmetric antenna layout, Euglena’s design appears optimized for robust yet adjustable energy capture.
What This Means for Sun-Powered Life
To a non-specialist, the key message is that evolution has many ways to solve the problem of turning sunlight into usable energy. Euglena gracilis, having acquired its chloroplast “second-hand,” did not simply copy the standard plant blueprint. Instead, it pared down the core, repositioned all antennas to one side, invented a new protein loop to anchor them, and deployed distinctive chlorophyll clusters and pigments to steer energy efficiently into its reaction center. This lineage-specific solution shows how secondary plastids can remodel the photosynthetic apparatus while preserving—and even enhancing—the basic task of harvesting light for life.
Citation: Huang, G., Dong, S., Li, L. et al. The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis. Nat Commun 17, 3429 (2026). https://doi.org/10.1038/s41467-026-70183-y
Keywords: Euglena gracilis, photosystem I, light-harvesting complexes, excitation energy transfer, cryo-electron microscopy