Clear Sky Science · pl
Architektura i szlaki transferu energii PSI–LHCI–LHCII w fototroficznym wiciowcu Euglena gracilis
Jak maleńki pływak zamienia światło słoneczne w energię
Jednokomórkowy glon Euglena gracilis to mikroskopijny pływak, który dobrze czuje się w stawach i kałużach. Podobnie jak rośliny, wykorzystuje światło słoneczne jako źródło energii, lecz robi to za pomocą aparatu fotosyntetycznego, który przeszedł nietypową drogę ewolucyjną. To badanie ujawnia ten mechanizm w skali atomowej, pokazując, jak Euglena przeprojektowała swój system zbierania światła, aby efektywnie i elastycznie wychwytywać energię po tym, jak miliony lat temu pozyskała chloroplast od innego glonu.
Specjalny silnik świetlny w zapożyczonym chloroplaście
W centrum fotosyntezy znajduje się Fotosystem I, kompleks białkowo-barwnikowy, który przekształca energię świetlną w przepływ elektronów potrzebny do budowy cukrów. U większości roślin i zielonych glonów jednostka ta otoczona jest sierpowatym łukiem białek antenowych zbierających światło. Euglena gracilis jest inna: jej chloroplast powstał przez pochłonięcie już fotosyntetyzującego zielonego glonu, dając tzw. plastydu wtórnego. Ten nietypowy pochodzenie pozostawiło Euglenę z charakterystycznymi pigmentami i składnikami białkowymi, a dotąd dokładny układ nadkompleksu Fotosystemu I nie był znany. Dzięki wysokorozdzielczej kriomikroskopii elektronowej autorzy odtworzyli trójwymiarową strukturę tego zespołu z rozdzielczością do 2,06 angstroma, ujawniając, jak Euglena przebudowała swój „silnik świetlny”.
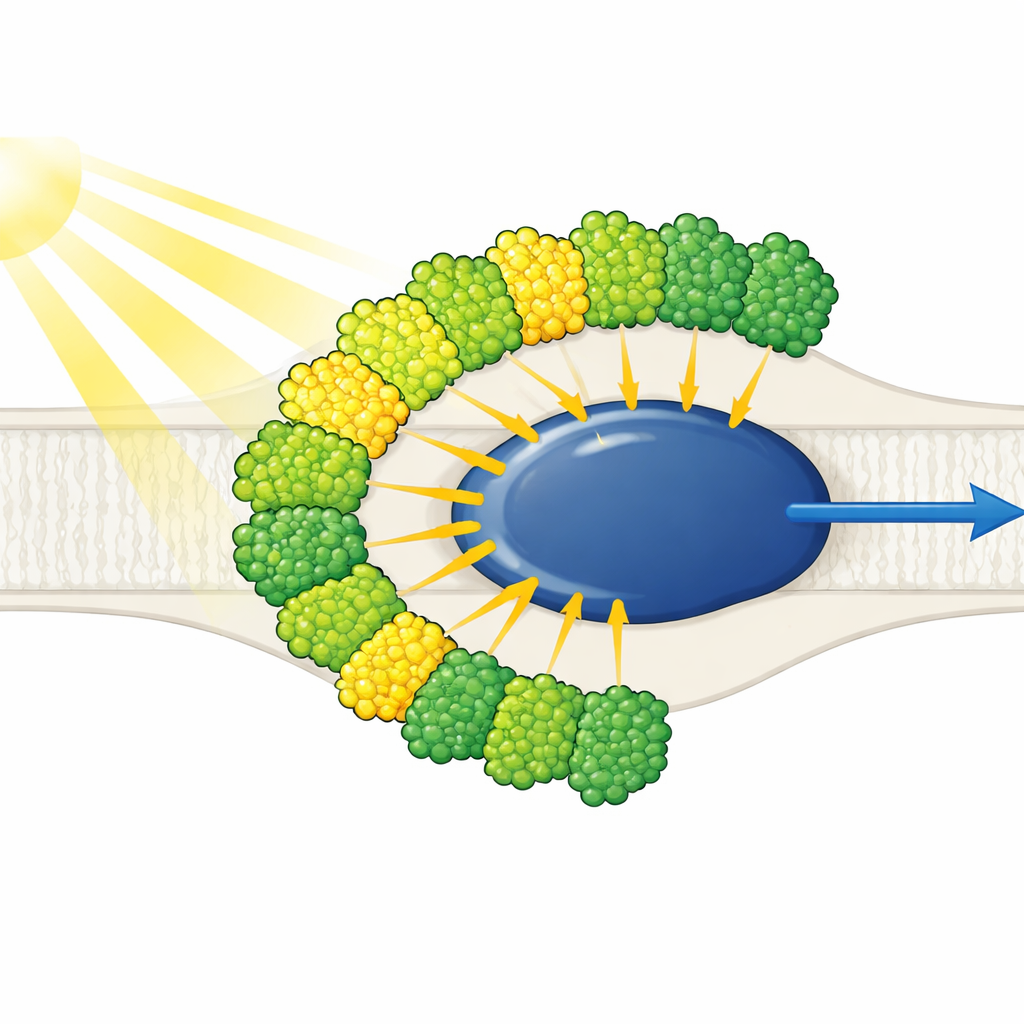
Anteny świetlne wszystkie po jednej stronie
Struktura ukazuje uproszczone jądro Fotosystemu I otoczone przez dziesięć anten zbierających światło — dziewięć jednego typu (LHCI) i jedną zwykle kojarzoną z Fotosystemem II (LHCII). Co godne uwagi, wszystkie dziesięć anten znajduje się tylko po jednej stronie jądra, w dwóch koncentrycznych pasach, zamiast tworzyć typowy dla roślin sierpowy łuk po przeciwnej stronie. Każda antena zewnętrzna paruje twarz do twarzy z wewnętrznym sąsiadem, tworząc pięć układów dimerów ułożonych warstwowo, które z kolei składają się w dwa tetramery. Anteny te zbudowane są z poliprotein, które są cięte na pojedyncze jednostki, a specyficzne krótkie motywy sekwencyjne działają jak molekularne rzepy spajające dimerów. Jedna wewnętrzna antena, nazwana LHC-3, wyłania się jako centralne ogniwo, łącząc dimery i pomagając przywiązać cały pas do jądra.
Nowa pętla białkowa zastępująca utracone elementy
W porównaniu z dobrze znanymi systemami roślinnymi jądro Fotosystemu I u Eugleny nie ma kilku małych białek kotwiczących, które zwykle pozycjonują anteny i umożliwiają ich dynamiczną reorganizację przy zmieniającym się świetle. Zamiast tego jeden z pozostałych podjednostek jądra, PsaD, niesie dodatkową pętlę o długości około 40 aminokwasów występującą wyłącznie u Eugleny. Ta elastyczna wstawka sięga w przestrzeń, gdzie normalnie znajdowałyby się brakujące białka, i tworzy bezpośrednie stabilizujące kontakty z LHC-3 i jego chlorofilami. Autorzy proponują, że ta unikatowa pętla ewoluowała równolegle z utratą brakujących podjednostek, przejmując ich rolę w mocowaniu anten do jądra i kształtując miejsce oraz sposób, w jaki napływająca energia świetlna wchodzi do maszyny.
Specjalne pigmenty i energetyczne autostrady
Ponad architekturą białek, rozmieszczenie cząsteczek chlorofilu determinuje, jak płynie energia świetlna. Kompleks Eugleny zawiera ponad 200 pigmentów, w tym chlorofile a i b oraz specjalne karotenoidy różniące się od tych u roślin lądowych. Dwa chlorofile, oznaczone a1020 i a1021 przez analogię do znanych miejsc, są unikatowe dla Eugleny i występują wyłącznie w antenach wewnętrznych. Leżą one w węzłach, gdzie spotykają się trzy anteny, tworząc skupiska pigmentów, które fizycznie łączą sąsiednie jednostki. Te skupiska, razem z blisko położonymi parami „czerwonych” chlorofili absorbujących światło o nieco niższej energii, tworzą wiele dróg przepływu wzbudzenia z pasa zewnętrznego do pasa wewnętrznego, a następnie do jądra. Obliczenia oparte na odległościach między centrami pigmentów wskazują, że energia zbierana w antenach zewnętrznych jest preferencyjnie kierowana do kilku bramkowych anten, zwłaszcza LHC-3 i jej odpowiednika LHC-9, zanim zostanie przekazana do centrum reakcji, gdzie zachodzi transfer elektronów.
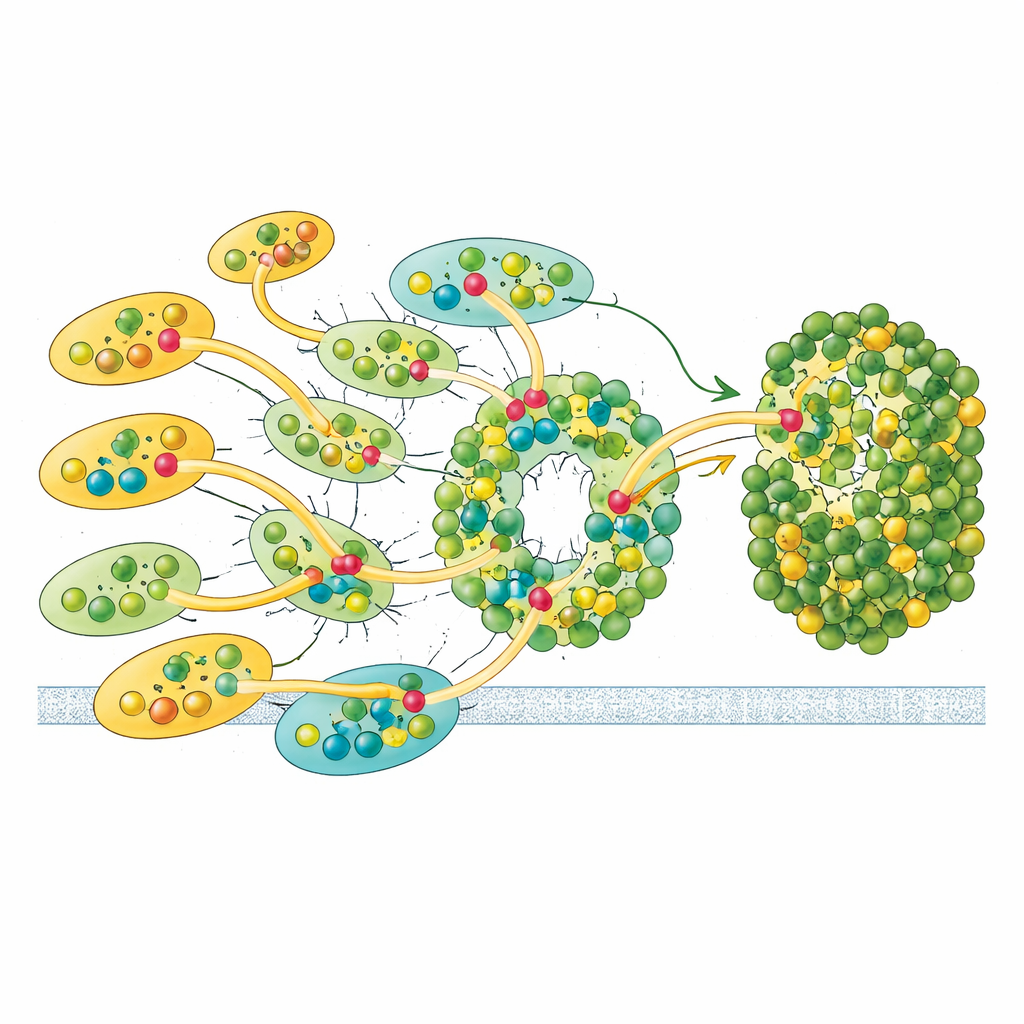
Elastyczne łącza do adaptacji przy zmiennym świetle
Nie wszystkie anteny są przymocowane równie mocno. Pojedyncza jednostka LHCII i jedna z zewnętrznych LHCI wykazują tylko częściową zajętość w danych kriomikroskopowych, co sugeruje, że mogą przyłączać się i odłączać łatwiej niż pozostałe. Ta zmienność wskazuje na funkcję regulacyjną: przez dodawanie lub usuwanie tych anten Euglena może dostrajać ilość światła docierającego do Fotosystemu I i korygować bilans energii dzielonej z Fotosystemem II, przypominając przejścia stanów znane z roślin, lecz realizując je innym rozwiązaniem strukturalnym. W połączeniu z unikatowymi pigmentami i asymetrycznym układem anten konstrukcja Eugleny wydaje się zoptymalizowana pod kątem solidnego, a zarazem regulowanego wychwytu energii.
Co to oznacza dla życia napędzanego słońcem
Dla osoby nietechnicznej kluczowe przesłanie jest takie, że ewolucja ma wiele sposobów na rozwiązanie problemu zamiany światła słonecznego w użyteczną energię. Euglena gracilis, pozyskawszy swój chloroplast „z drugiej ręki”, nie skopiowała po prostu standardowego planu roślinnego. Zamiast tego uprościła jądro, przesunęła wszystkie anteny na jedną stronę, wymyśliła nową pętlę białkową do ich zakotwiczenia i zastosowała charakterystyczne skupiska chlorofili oraz pigmenty, które efektywnie kierują energię do centrum reakcji. To specyficzne dla linii rozwią- zanie pokazuje, jak plastydy wtórne mogą przeprojektować aparat fotosyntetyczny, zachowując — a nawet polepszając — podstawowe zadanie zbierania światła dla życia.
Cytowanie: Huang, G., Dong, S., Li, L. et al. The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis. Nat Commun 17, 3429 (2026). https://doi.org/10.1038/s41467-026-70183-y
Słowa kluczowe: Euglena gracilis, photosystem I, kompleksy zbierające światło, transfer energii wzbudzenia, kriomikroskopia elektronowa