Clear Sky Science · de
Die Architektur und Energieübertragungswege von PSI–LHCI–LHCII im phototrophen Geißeltierchen Euglena gracilis
Wie ein winziger Schwimmer Sonnenlicht in Energie verwandelt
Die einzellige Alge Euglena gracilis ist ein mikroskopischer Schwimmer, der in Teichen und Pfützen gedeiht. Wie Pflanzen nutzt sie Sonnenlicht, um Leben zu ermöglichen, doch tut sie dies mit einer photosynthetischen Maschine, die einen ungewöhnlichen evolutionären Weg genommen hat. Diese Studie zeigt diese Maschine in atomaren Details und legt dar, wie Euglena sein lichtsammelndes System nach dem Erwerb seines Chloroplasten von einer anderen Alge vor Millionen von Jahren effizient und flexibel umgebaut hat.
Ein spezieller Lichtmotor in einem geborgten Chloroplasten
Im Zentrum der Photosynthese steht Photosystem I, ein Protein‑Pigment‑Komplex, der Lichtenergie in den Elektronenfluss umwandelt, der zum Aufbau von Zuckern nötig ist. Bei den meisten Pflanzen und Grünalgen ist diese Kerneinheit von einem Halbmond aus Antennenproteinen umgeben, die Licht sammeln. Euglena gracilis unterscheidet sich: Sein Chloroplast entstand durch die Aufnahme einer bereits photosynthetischen Grünalge und ist damit ein sogenannter sekundärer Plastid. Dieser ungewöhnliche Ursprung hinterließ bei Euglena charakteristische Pigmente und Proteinbestandteile, und bis jetzt war die genaue Anordnung seines Photosystem‑I‑Superkomplexes unbekannt. Mittels hochauflösender Kryo‑Elektronenmikroskopie lösten die Autoren die dreidimensionale Struktur dieser Anordnung bis zu 2,06 Å auf und zeigen, wie Euglena seinen Lichtmotor umgebaut hat.
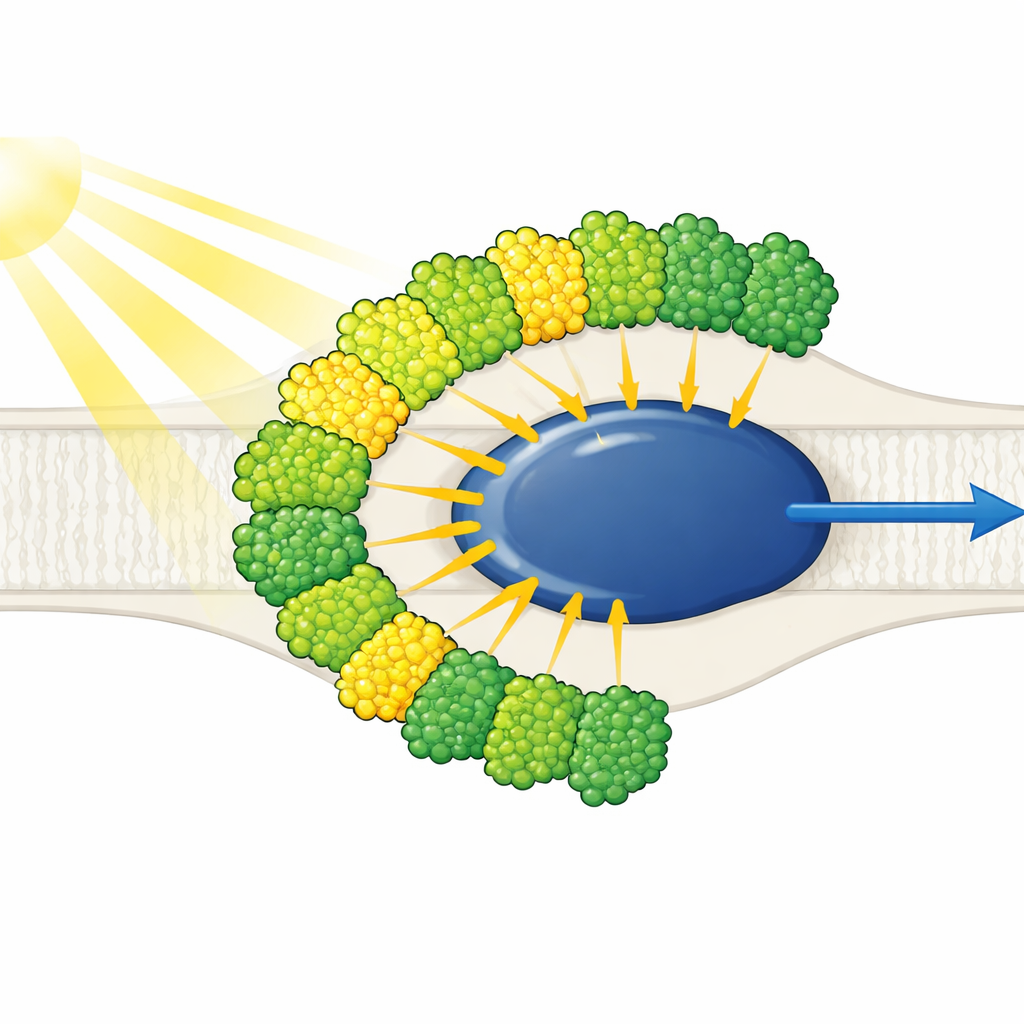
Lichtantennen alle auf einer Seite
Die Struktur zeigt einen reduziert wirkenden Photosystem‑I‑Kern, umgeben von zehn lichtsammelnden Antennen — neun eines Typs (LHCI) und eine, die normalerweise mit Photosystem II assoziiert wird (LHCII). Auffällig ist, dass alle zehn Antennen nur auf einer Seite des Kerns sitzen, in zwei konzentrischen Gürtelreihen, statt die für Pflanzen typische halbmondförmige Anordnung auf der gegenüberliegenden Seite zu bilden. Jede äußere Antenne steht paarweise gegenüber einer inneren Nachbarin, wodurch fünf geschichtete Dimere entstehen, die sich wiederum zu zwei Tetrameren zusammenlagern. Diese Antennen bestehen aus Polyproteinen, die in einzelne Einheiten geschnitten werden, und spezifische kurze Sequenzmotive wirken wie molekulares Klettband, das die Dimere zusammenhält. Eine innere Antenne, LHC‑3 genannt, tritt als zentrales Verbindungselement hervor, verbindet Dimere und hilft, den gesamten Gürtel mit dem Kern zu verankern.
Eine neue Proteinschleife ersetzt verlorene Teile
Im Vergleich zu vertrauten Pflanzensystemen fehlt im Photosystem‑I‑Kern von Euglena eine Reihe kleiner Ankerproteine, die normalerweise Antennen positionieren und die dynamische Umorganisation bei wechselndem Licht ermöglichen. Stattdessen trägt eine der verbleibenden Kernuntereinheiten, PsaD, eine zusätzliche Schleife von etwa 40 Aminosäuren, die nur in Euglena vorkommt. Diese flexible Einfügung reicht in den Raum hinein, in dem die fehlenden Proteine sonst sitzen würden, und bildet direkte stabilisierende Kontakte mit LHC‑3 und dessen Chlorophyllen. Die Autoren stellen die Hypothese auf, dass diese einzigartige Schleife gleichzeitig mit dem Verlust der fehlenden Untereinheiten evolvierte und deren Rolle beim Befestigen der Antennen am Kern übernahm sowie die Positionen und Wege steuerte, über die eintreffende Lichtenergie in die Maschine gelangt.
Maßgeschneiderte Pigmente und Energieautobahnen
Über die Proteinarchitektur hinaus bestimmt die Positionierung der Chlorophyllmoleküle, wie Lichtenergie fließt. Der Euglena‑Komplex trägt über 200 Pigmente, darunter Chlorophylle a und b sowie spezielle Carotinoide, die sich von denen höherer Pflanzen unterscheiden. Zwei Chlorophylle, analog zu bekannten Stellen als a1020 und a1021 bezeichnet, sind uniquely für Euglena und erscheinen nur in den inneren Antennen. Sie sitzen an Knotenpunkten, an denen drei Antennen zusammenlaufen, und bilden Pigmentcluster, die benachbarte Einheiten physikalisch überbrücken. Diese Cluster, zusammen mit eng gepaarten „roten“ Chlorophyllen, die etwas energieärmer absorbieren, schaffen mehrere Routen, auf denen Anregungsenergie vom äußeren Gürtel in den inneren Gürtel und dann in den Kern geleitet wird. Rechnungen, die auf Abständen zwischen Pigmentzentren basieren, zeigen, dass die in den äußeren Antennen gesammelte Energie bevorzugt über wenige Tor‑Antennen kanalisiert wird — insbesondere LHC‑3 und sein Gegenstück LHC‑9 — bevor sie an das Reaktionszentrum übergeben wird, wo der Elektronentransfer stattfindet.
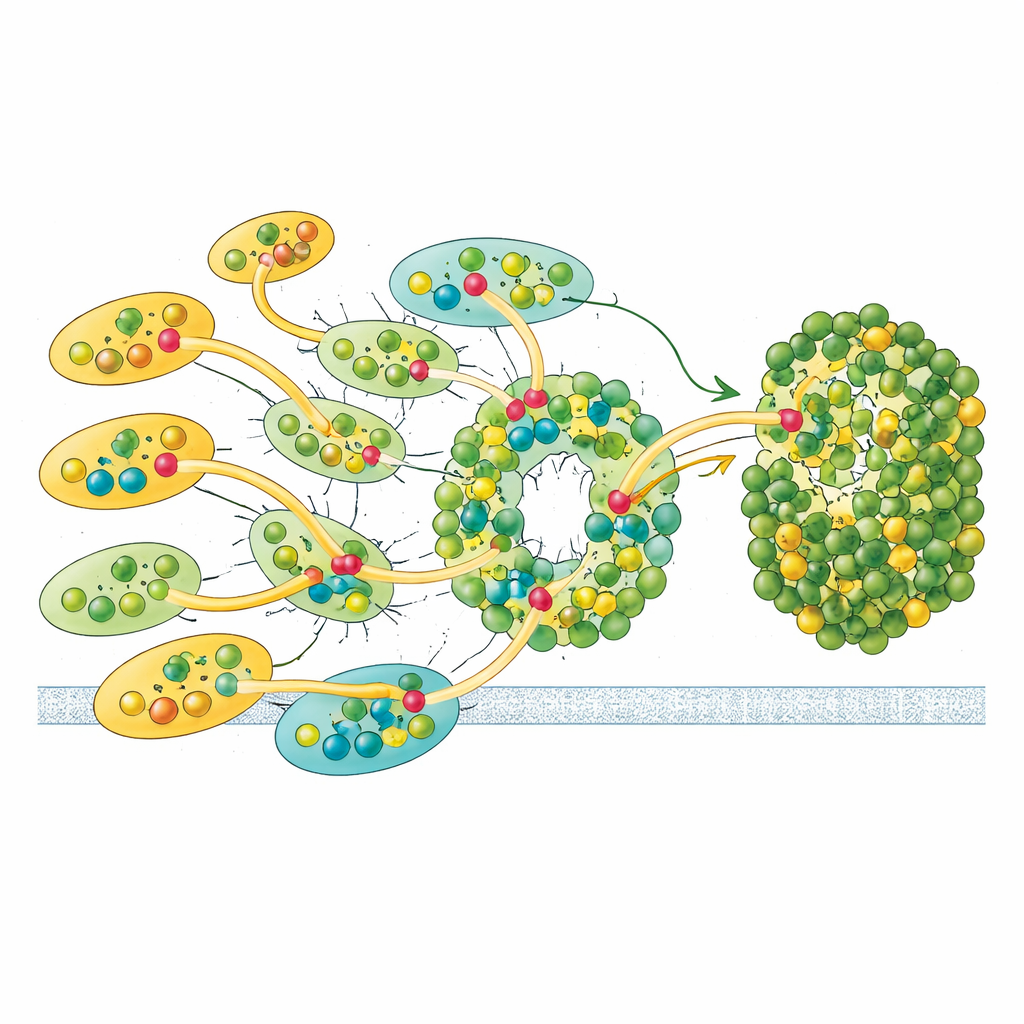
Flexiblen Verbindungen zur Anpassung an wechselndes Licht
Nicht alle Antennen sind gleich fest verankert. Die einzelne LHCII‑Einheit und eine äußere LHCI zeigen in den Kryo‑EM‑Daten nur partielle Besetzung, was darauf hindeutet, dass sie sich leichter an‑ und abkoppeln können als die anderen. Diese Labilität deutet auf eine Regulationsfunktion hin: Durch Hinzufügen oder Entfernen dieser Antennen kann Euglena möglicherweise steuern, wie viel Licht Photosystem I erreicht, und das Verhältnis der Energieverteilung zu Photosystem II anpassen — ein Echo der in Pflanzen bekannten Zustandstransitionen, hier jedoch über eine andere strukturelle Lösung. Zusammen mit seinen einzigartigen Pigmenten und der asymmetrischen Antennenanordnung scheint Euglenas Design auf robuste, dabei anpassbare Energieerfassung optimiert zu sein.
Was das für sonnenbetriebene Lebensformen bedeutet
Für Nicht‑Spezialisten lautet die Kernbotschaft, dass die Evolution viele Wege kennt, das Problem zu lösen, Sonnenlicht in nutzbare Energie zu verwandeln. Euglena gracilis, das seinen Chloroplasten „zweithändig“ erhalten hat, hat nicht einfach den Standardplan der Pflanzen kopiert. Stattdessen hat es den Kern reduziert, alle Antennen auf eine Seite verlagert, eine neue Proteinschleife erfunden, um sie zu verankern, und charakteristische Chlorophyllcluster sowie Pigmente eingesetzt, um Energie effizient in sein Reaktionszentrum zu lenken. Diese linien‑spezifische Lösung zeigt, wie sekundäre Plastiden das photosynthetische System umgestalten können, während sie die grundsätzliche Aufgabe des Lichtsammelns für das Leben bewahren — und in manchen Aspekten sogar verbessern.
Zitation: Huang, G., Dong, S., Li, L. et al. The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis. Nat Commun 17, 3429 (2026). https://doi.org/10.1038/s41467-026-70183-y
Schlüsselwörter: Euglena gracilis, Photosystem I, Lichtsammlungs-Komplexe, Anregungsenergieübertragung, Kryo-Elektronenmikroskopie