Clear Sky Science · ru
Архитектура и пути передачи энергии PSI–LHCI–LHCII в фототрофном жгутиконосце Euglena gracilis
Как крошечный пловец превращает солнечный свет в энергию
Одноклеточная водоросль Euglena gracilis — микроскопический пловец, процветающий в прудах и лужах. Подобно растениям, она использует солнечный свет для поддержания жизни, но делает это с помощью фотосинтетического аппарата, прошедшего необычный эволюционный путь. В этом исследовании машина показана с атомной детальностью: раскрыто, как Euglena перенастроила свою систему сбора света, чтобы эффективно и гибко захватывать энергию после того, как миллионы лет назад получила хлоропласт от другой водоросли.
Особый световой двигатель в «заимствованном» хлоропласте
В основе фотосинтеза лежит фотосистема I — белково-пигментный комплекс, превращающий энергию света в поток электронов, необходимый для синтеза сахаров. У большинства растений и зелёных водорослей этот ядро окружено полумесяцем антенных белков, собирающих свет. Euglena gracilis отличается: её хлоропласт возник в результате поглощения уже фотосинтезирующей зелёной водоросли, то есть представляет собой вторичный пласти́д. Такое необычное происхождение оставило у Euglena характерные пигменты и белковые компоненты, и до сих пор точная организация суперкомплекса фотосистемы I была неизвестна. С помощью высокоразрешающей крио-ЭМ авторы определили трёхмерную структуру этой сборки с разрешением до 2,06 ангстрем, показав, как Euglena перестроила свой световой двигатель.
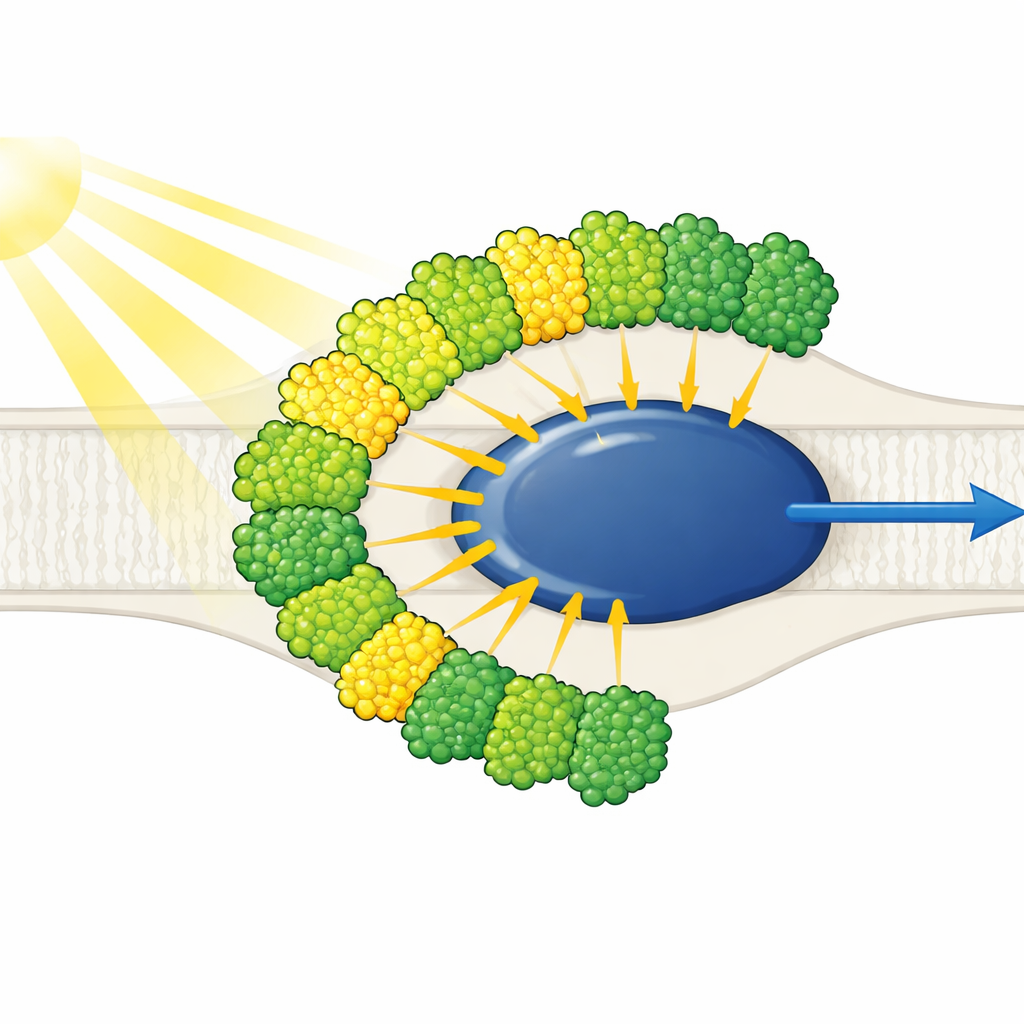
Антенны света — все с одной стороны
Структура показывает упрощённое ядро фотосистемы I, окружённое десятью антеннами сбора света — девятью одного типа (LHCI) и одной, обычно связанной с фотосистемой II (LHCII). Примечательно, что все десять антенн расположены с одной стороны ядра, в двух концентрических поясах, а не в типичном для растений полумесяце на противоположной стороне. Каждая внешняя антенна стоит лицом к внутренней, образуя пять сцепленных димеров, которые затем собираются в два тетрамера. Эти антенны состоят из полипротеинов, разрезаемых на отдельные единицы, а короткие специфические последовательности работают как молекулярный «велкро», удерживая димеры вместе. Одна из внутренних антенн, названная LHC-3, выступает как центральный узел, соединяющий димеры и помогая привязать весь пояс к ядру.
Новая белковая петля вместо утраченных частей
По сравнению со знакомыми растительными системами ядро фотосистемы I у Euglena лишено нескольких небольших якорных белков, которые обычно позиционируют антенны и позволяют динамическую перестройку при изменении освещённости. Вместо этого один из оставшихся субъединиц ядра, PsaD, несёт дополнительную петлю приблизительно из 40 аминокислот, встречающуюся только у Euglena. Эта гибкая вставка проникает в пространство, где обычно располагались утраченные белки, и образует прямые стабилизирующие контакты с LHC-3 и его хлорофилл‑молекулами. Авторы предполагают, что эта уникальная петля эволюционировала параллельно с потерей указанных субъединиц, взяв на себя их роль в закреплении антенн к ядру и формировании путей входа поглощённой энергии в машину.
Специальные пигменты и энергетические магистрали
Помимо архитектуры белков, расположение молекул хлорофилла определяет, как течёт световая энергия. Комплекс Euglena содержит более 200 пигментов, включая хлорофиллы a и b и особые каротиноиды, отличающиеся от наземных растений. Два хлорофилла, обозначенные как a1020 и a1021 по аналогии с известными сайтами, уникальны для Euglena и обнаруживаются только во внутренних антеннах. Они расположены в узлах, где сходятся три антенны, образуя пигментные кластеры, физически связывающие соседние единицы. Эти кластеры вместе с плотно расположенными «красными» парами хлорофиллов, поглощающими немного более низкоэнергетический свет, создают множественные маршруты для перемещения возбуждения от внешнего пояса к внутреннему и далее в ядро. Расчёты на основе расстояний между центрами пигментов показывают, что энергия, собранная внешними антеннами, направляется преимущественно в несколько «воротных» антенн, особенно в LHC-3 и её партнёр LHC-9, прежде чем быть переданной в реакционный центр, где происходит перенос электронов.
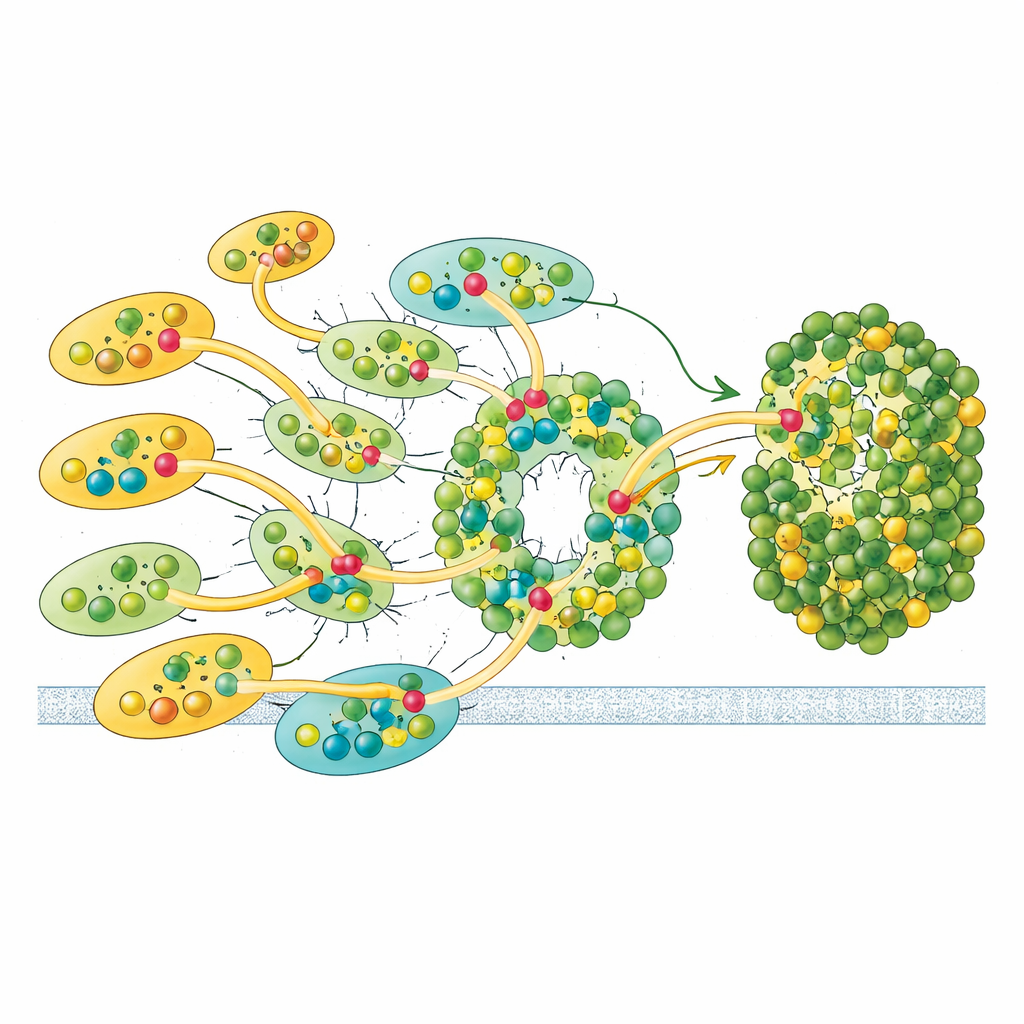
Гибкие связи для адаптации к изменяющемуся свету
Не все антенны прикреплены одинаково прочно. Единственная единица LHCII и одна внешняя LHCI показывают лишь частичную заполненность в данных крио-ЭМ, что указывает на способность этих модулей присоединяться и отсоединяться легче, чем другие. Эта подвижность намекает на регуляторную функцию: добавляя или удаляя эти антенны, Euglena может настраивать, сколько света достигает фотосистемы I, и корректировать баланс энергии, разделяемой с фотосистемой II, напоминая переходы состояний, известные у растений, но реализуя их иным структурным способом. В сочетании с уникальными пигментами и асимметричным расположением антенн конструкция Euglena выглядит оптимизированной для надёжного, но при этом регулируемого захвата энергии.
Что это значит для жизни, питаемой солнцем
Для неспециалиста главный вывод таков: эволюция предлагает множество решений проблемы превращения солнечного света в пригодную энергию. Euglena gracilis, получившая свой хлоропласт «со вторых рук», не скопировала стандартную растительную схему. Вместо этого она упростила ядро, переместила все антенны на одну сторону, создала новую белковую петлю для их закрепления и задействовала специфические кластеры хлорофиллов и пигменты, чтобы эффективно направлять энергию в реакционный центр. Это видоспецифическое решение показывает, как вторичные пластида способны перестраивать фотосинтетический аппарат, сохраняя — а порой и улучшая — основную задачу сбора света для поддержания жизни.
Цитирование: Huang, G., Dong, S., Li, L. et al. The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis. Nat Commun 17, 3429 (2026). https://doi.org/10.1038/s41467-026-70183-y
Ключевые слова: Euglena gracilis, фотосистема I, комплексы сбора света, передача возбуждённой энергии, криоэлектронная микроскопия