Clear Sky Science · ja
光栄沢性鞭毛虫Euglena gracilisにおけるPSI–LHCI–LHCIIの構造とエネルギー伝達経路
小さな泳ぎ手が太陽光をエネルギーに変えるしくみ
単細胞藻類Euglena gracilisは池や水たまりで繁栄する顕微鏡的な遊泳者です。植物と同様に太陽光を利用して生命活動を維持しますが、その光合成機構は特異な進化経路をたどってきました。本研究はその機構を原子分解能で明らかにし、ユーグレナが別の藻類から葉緑体を獲得した何百万年後に、光捕集系を効率的かつ柔軟に動作するよう再配線した様子を示します。
借り物の葉緑体に組み込まれた特別な光のエンジン
光合成の中核にあるのは光エネルギーを電子の流れに変換して糖の合成を支える光合成系I(Photosystem I)というタンパク質–色素複合体です。多くの植物や緑藻では、この中核ユニットは光を集めるアンテナ蛋白で三日月状に取り囲まれています。Euglena gracilisは異なります:その葉緑体は既に光合成能を持つ緑藻を取り込むことで成立したいわゆる二次プラスチドに由来します。この特殊な起源は独自の色素やタンパク質成分をもたらし、これまでEuglenaのPhotosystem I超複合体の正確な配置は不明でした。高解像度クライオ電子顕微鏡を用いて、著者らはこのアセンブリの三次元構造を2.06オングストロームまで解決し、Euglenaがどのように光のエンジンを再構築したかを明らかにしました。
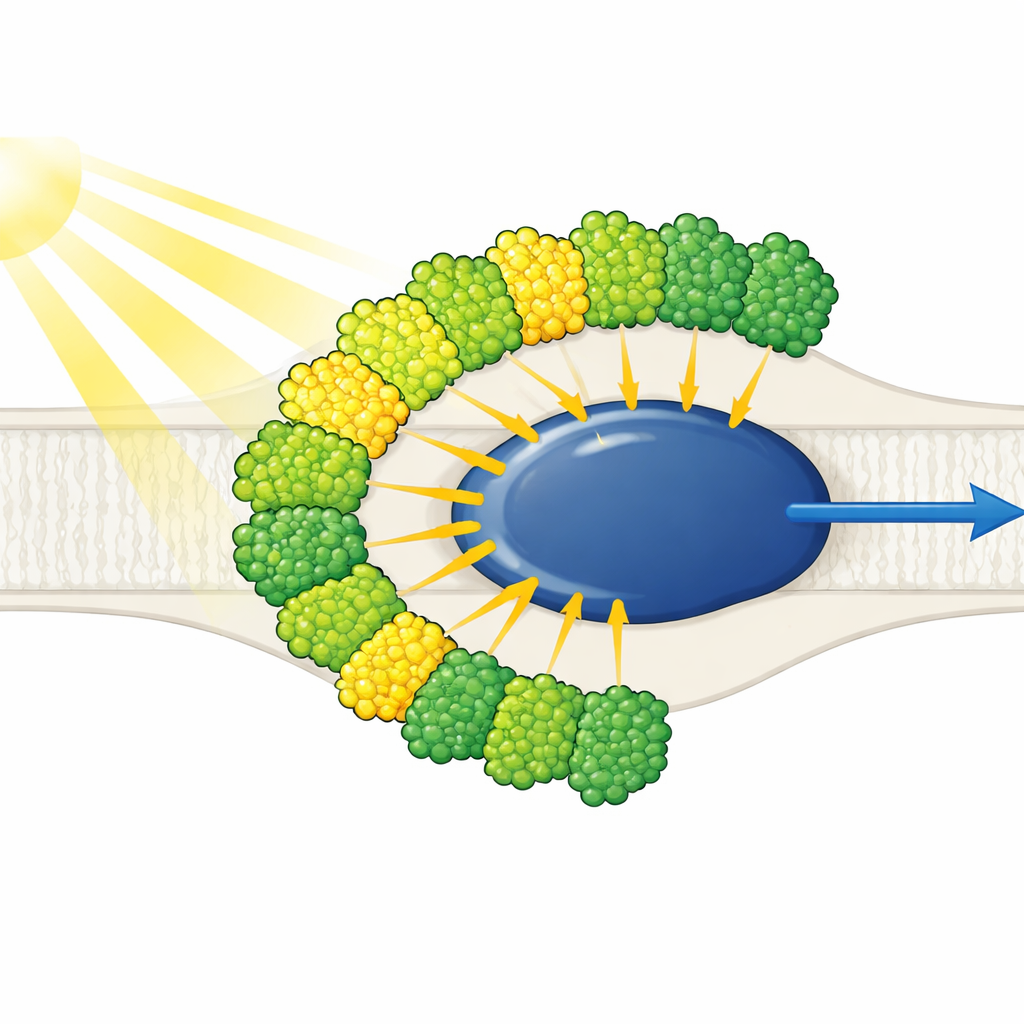
アンテナはすべて片側に集中
構造解析は、簡素化されたPhotosystem Iコアが十個の光捕集アンテナに囲まれていることを示します――九個はLHCI型、もう一つは通常Photosystem IIに関連づけられるLHCII型です。注目すべきは、これら十個のアンテナがコアの片側にのみ配置され、植物で見られるような反対側の三日月形ではなく、二つの同心ベルトを形成している点です。外側の各アンテナは内側の近接アンテナと対向しており、五つの積み重なったジマーを作り、さらにこれらが二つの四量体へと集合しています。これらのアンテナはポリタンパク質から切り出されて個々のユニットとなり、特定の短い配列モチーフが分子的な面ファスナーのようにジマーを結び付けています。内側の一つのアンテナ、LHC-3は中心的なハブとして浮かび上がり、ジマーをつなぎ合わせることで全体のベルトをコアに固定する役割を担っています。
失われた部位の代わりになる新しいタンパク質ループ
馴染みのある植物系と比べると、EuglenaのPhotosystem Iコアはアンテナの位置決めや光条件の変化に応じた動的再編成を助けるいくつかの小さなアンカータンパク質を欠いています。その代わりに残存するコアサブユニットの一つであるPsaDが、Euglenaに特有の約40アミノ酸から成る余分なループを持っています。この柔軟な挿入領域は通常それら欠損部分が位置する空間へ届き、LHC-3とそのクロロフィルと直接的な安定化相互作用を形成します。著者らは、この独特のループが欠失したサブユニット群とともに進化し、アンテナをコアに固定する役割を引き継ぎ、入射した光エネルギーがどこからどのように機械へ入るかを形づくっていると提案します。
特注の色素とエネルギーの高速道路
タンパク質の構造を越えて、クロロフィル分子の配置が光エネルギーの流れを決定します。Euglena複合体は200個を超える色素をもち、クロロフィルaとbに加え、陸上植物とは異なる特異なカロテノイドを含みます。既知の部位に倣ってa1020およびa1021と名付けられた二つのクロロフィルはEuglenaに固有で、内側アンテナにのみ見られます。これらは三つのアンテナが合流する接合部に位置し、隣接ユニットを物理的につなぐ色素クラスターを形成しています。これらのクラスターは、わずかに低エネルギーを吸収する“赤”クロロフィルペアの密接な配列とともに、外側ベルトから内側ベルト、そしてコアへと励起を運ぶ複数の経路を生み出します。色素中心間の距離に基づく計算は、外側アンテナで集められたエネルギーが少数のゲートウェイ的アンテナ、特にLHC-3とその対応者LHC-9へ優先的に注ぎ込まれ、そこで反応中心へ電子移動が起こる方向へ受け渡されることを示しています。
変わりゆく光に適応する柔軟な連結
すべてのアンテナが同じようにがっちり固定されているわけではありません。孤立したLHCIIユニットと一つの外側LHCIはクライオEMデータで部分的な占有率しか示さず、これらは他のアンテナよりも着脱しやすい可能性が示唆されます。この可変性は調節機能を示唆します:これらのアンテナを付け外しすることで、EuglenaはPhotosystem Iに届く光量を調整し、Photosystem IIとのエネルギー分配のバランスを変えることができるかもしれません。これは植物で知られるステート遷移と同様の効果を反映しますが、異なる構造的解決策を用いています。独自の色素と非対称なアンテナ配置と合わせて、Euglenaの設計は堅牢でありながら調整可能な光捕集に最適化されているようです。
太陽エネルギー駆動の生命にとっての意義
専門外の読者にとって重要な点は、太陽光を実用的なエネルギーに変換する問題に対して進化が多様な解を提供することです。二次プラスチドを“間借り”したEuglena gracilisは、標準的な植物の設計図を単にコピーしたわけではありません。むしろコアを簡素化し、すべてのアンテナを片側に再配置し、新しいタンパク質ループを発明してそれらを固定し、独特のクロロフィルクラスターと色素を配置することで、反応中心へ効率的にエネルギーを導くように構築しました。この系統特異的な解決策は、二次プラスチドが光合成装置を再構築しつつ、光を生命のために捕らえる基本的な任務を保持し、場合によっては強化することを示しています。
引用: Huang, G., Dong, S., Li, L. et al. The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis. Nat Commun 17, 3429 (2026). https://doi.org/10.1038/s41467-026-70183-y
キーワード: Euglena gracilis, 光合成系I, 光捕集複合体, 励起エネルギー移動, クライオ電子顕微鏡法