Clear Sky Science · tr
Euglena gracilis'te PSI–LHCI–LHCII mimarisı ve enerji aktarım yolları
Minik Bir Yüzücünün Güneş Işığını Güce Çevirme Yolu
Tek hücreli alg Euglena gracilis, gölet ve su birikintilerinde yaşayan mikroskobik bir yüzücüdür. Bitkiler gibi yaşamı güneş ışığıyla besler; ancak bunu alışılmadık bir evrimsel yol izleyen bir fotosentetik makineyle yapar. Bu çalışma, o makineyi atomik ayrıntıda ortaya koyarak, Euglena'nın kloroplastunu milyonlarca yıl önce başka bir algden edindikten sonra ışık toplama sistemini enerji yakalama açısından verimli ve esnek biçimde nasıl yeniden düzenlediğini gösteriyor.
Ödünç Alınmış Bir Kloroplastta Özel Bir Işık Motoru
Fotosentezin merkezinde, ışık enerjisini şeker üretimi için gerekli elektron akışına çeviren protein–pigment kompleksi Fotosistem I bulunur. Çoğu bitki ve yeşil algde bu temel birimin çevresinde ışığı toplayan bir çember şeklinde anten proteinleri dizilir. Euglena gracilis farklıdır: kloroplastu, zaten fotosentetik bir yeşil algi yutarak edinilmiş, yani biyologların dediği gibi ikincil bir plastidlere sahiptir. Bu sıra dışı köken Euglena'ya ayırt edici pigmentler ve protein bileşenleri kazandırdı; bugüne kadar Fotosistem I süperkümelenmesinin tam düzeni bilinmiyordu. Yüksek çözünürlüklü kriyo-elektron mikroskobu kullanarak yazarlar bu yapının üç boyutlu yapısını 2.06 angström çözünürlüğe kadar çözdü ve Euglena'nın ışık motorunu nasıl yeniden kurduğunu ortaya koydu.
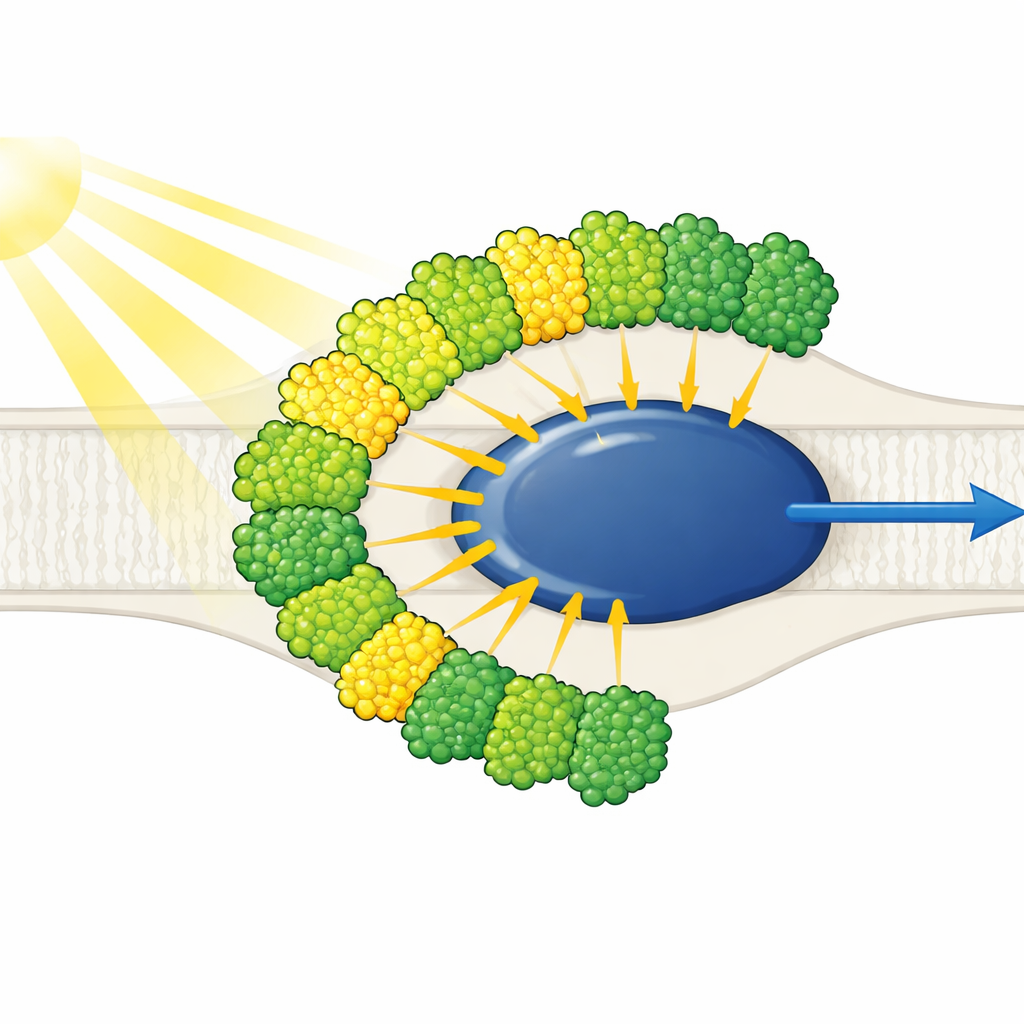
Tüm Işık Antenleri Tek Tarafa Dizilmiş
Yapı, sadeleştirilmiş bir Fotosistem I çekirdeğinin etrafında on ışık-toplayıcı anten—dokuz tanesi bir tür (LHCI) ve bir tanesi genellikle Fotosistem II ile ilişkilendirilen (LHCII)—ile sarılı olduğunu gösterir. Dikkat çekici şekilde tüm on anten yalnızca çekirdeğin tek bir tarafında, bitişik iki konsantrik kuşak halinde yer alır; tipik bitkilere özgü hilal formunu ters tarafta oluşturmazlar. Her dış anten, yüz yüze bir iç komşuyla eşleşerek beş istiflenmiş dimer oluşturur; bu dimerler de iki tetramer halinde bir araya gelir. Bu antenler, bireysel birimlere kesilen poli-proteinlerden oluşur ve belirli kısa dizi motifleri dimerleri bir arada tutan moleküler Velcro gibi iş görür. LHC-3 adını taşıyan bir iç anten, dimerleri birbirine bağlayan ve tüm kuşağı çekirdeğe bağlamaya yardımcı olan merkezi bir merkez olarak öne çıkar.
Kayıp Parçaların Yerini Alan Yeni Bir Protein Döngüsü
Tanıdık bitki sistemleriyle karşılaştırıldığında, Euglena'nın Fotosistem I çekirdeği, antenleri konumlandırmaya ve değişen ışığa göre dinamik yeniden örgütlenmeye yardımcı olan birkaç küçük ankraj proteininden yoksundur. Bunun yerine, geriye kalan çekirdek altbirimlerinden biri olan PsaD, yalnızca Euglena'da bulunan yaklaşık 40 amino asitlik ekstra bir halka taşıyor. Bu esnek ekleme, eksik proteinlerin normalde oturduğu alana uzanır ve LHC-3 ile onun klorofillerine doğrudan dengeleyici temaslar kurar. Yazarlar, bu benzersiz halkanın, kaybolan altbirimlerle birlikte evrimleşerek antenleri çekirdeğe sabitleme görevini devraldığını ve gelen ışık enerjisinin makineye nereden ve nasıl girdiğini şekillendirdiğini öne sürüyorlar.
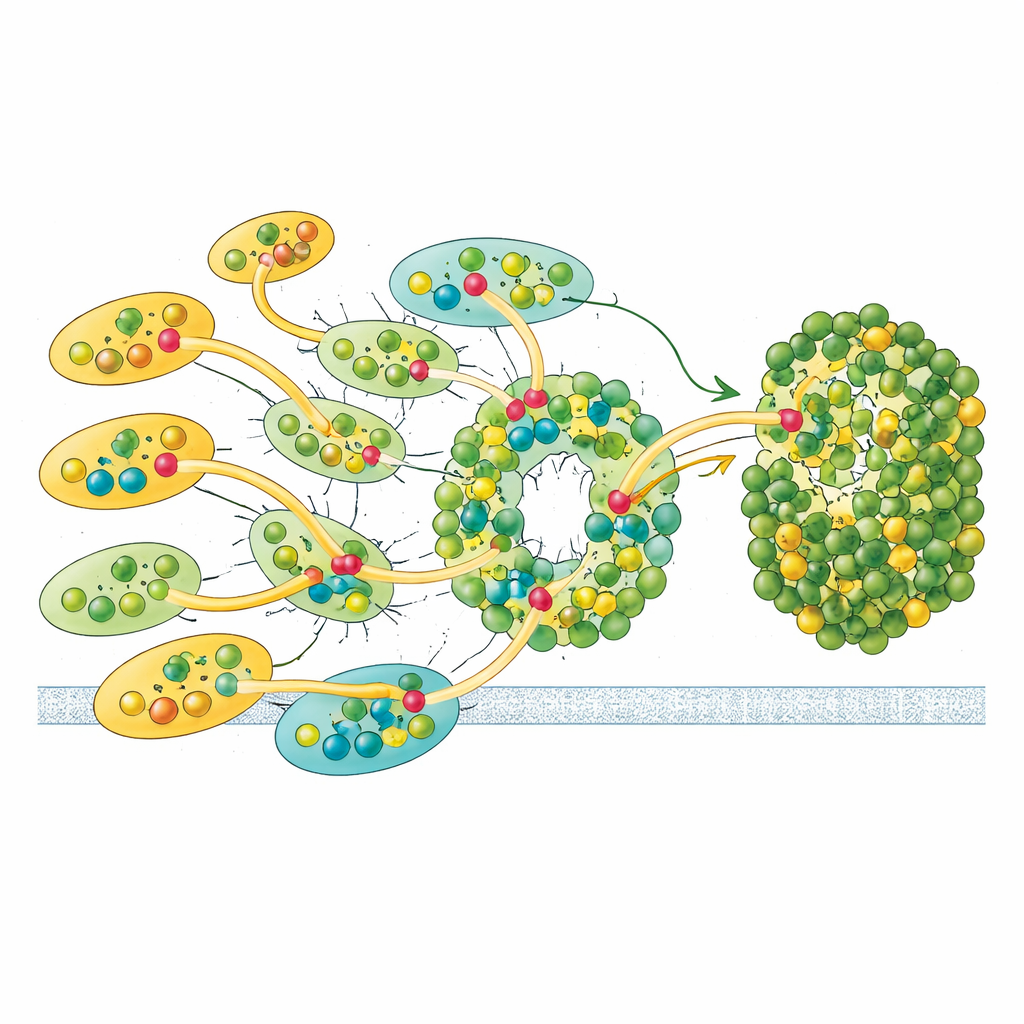
Özel Pigmentler ve Enerji Otoyolları
Protein mimarisinin ötesinde, klorofil moleküllerinin konumlanma biçimi ışık enerjisinin akışını belirler. Euglena kompleksi, klorofil a ve b ile kara bitkilerdekinden farklı özel karotenoidler de dahil olmak üzere 200'ün üzerinde pigment taşır. a1020 ve a1021 olarak adlandırılan iki klorofil, bilinen yerlere benzerlikleriyle Euglena'ya özgüdür ve yalnızca iç antenlerde bulunur. Bu klorofiller, üç antenin kesiştiği noktalarda yer alır ve komşu birimleri fiziksel olarak bağlayan pigment kümeleri oluşturur. Bu kümeler, ayrıca biraz daha düşük enerjili ışığı absorbe eden yakından yerleşmiş "kırmızı" klorofil çiftleriyle birlikte, uyarımın dış kuşaktan iç kuşağa ve oradan çekirdeğe gitmesi için birden çok yol oluşturur. Pigment merkezleri arasındaki mesafelere dayalı hesaplamalar, dış antenlerde toplanan enerjinin öncelikle birkaç kapı antenine—özellikle LHC-3 ve onun karşılığı LHC-9—a yönlendirildiğini ve buradan elektron transferinin gerçekleştiği reaksiyon merkezine aktarıldığını gösterir.
Değişen Işığa Uyum İçin Esnek Bağlar
Tüm antenler eşit derecede sıkı şekilde tutturulmamıştır. Tekil LHCII birimi ve bir dış LHCI, kriyo-EM verilerinde yalnızca kısmi işgal gösterir; bu da bunların diğerlerine göre daha kolay takılıp çıkarılabildiğini düşündürür. Bu hareketlilik düzenleyici bir özellik olabileceğine işaret eder: Euglena, bu antenleri ekleyip çıkararak Fotosistem I'e ulaşan ışık miktarını ayarlayabilir ve Fotosistem II ile paylaşılan enerjinin dengesini değiştirebilir—bu, bitkilerde bilinen durum geçişlerini andırır ama farklı bir yapısal çözüm kullanır. Benzersiz pigmentleri ve asimetrik anten düzeniyle birlikte Euglena'nın tasarımı, dayanıklı ancak ayarlanabilir enerji toplama için optimize edilmiş görünür.
Güneşle Çalışan Yaşam İçin Ne Anlama Geliyor
Uzman olmayan bir okur için ana mesaj şudur: Evrim, güneş ışığını kullanılabilir enerjiye dönüştürme sorununu çözmek için birçok yol sunar. Kloroplastunu "ikinci el" edinmiş olan Euglena gracilis, standart bitki taslağını basitçe kopyalamadı. Bunun yerine çekirdeği sadeleştirdi, tüm antenleri tek tarafa yerleştirdi, onları sabitlemek için yeni bir protein halkası geliştirdi ve enerjiyi reaksiyon merkezine verimli biçimde yönlendirmek üzere ayırt edici klorofil kümeleri ve pigmentler kullandı. Bu türden soy hattına özgü çözümler, ikincil plastidlerin fotosentetik aparatı yeniden şekillendirebileceğini ve temel ışık toplama görevini korurken hatta geliştirebileceğini gösterir.
Atıf: Huang, G., Dong, S., Li, L. et al. The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis. Nat Commun 17, 3429 (2026). https://doi.org/10.1038/s41467-026-70183-y
Anahtar kelimeler: Euglena gracilis, fotosistem I, ışık-toplayıcı kompleksler, uyarılmış enerji transferi, kriyofreze elektron mikroskobu