Clear Sky Science · es
La arquitectura y las vías de transferencia de energía de PSI–LHCI–LHCII en el flagelado fotótrofo Euglena gracilis
Cómo un pequeño nadador convierte la luz solar en energía
El alga unicelular Euglena gracilis es un nadador microscópico que prospera en estanques y charcos. Como las plantas, utiliza la luz solar para alimentar sus procesos vitales, pero lo hace con una maquinaria fotosintética que siguió un camino evolutivo poco habitual. Este estudio revela esa maquinaria con detalle atómico, mostrando cómo Euglena ha reconfigurado su sistema colector de luz para captar la energía de forma eficiente y flexible tras haber adquirido su cloroplasto de otro alga hace millones de años.
Un motor de luz especial en un cloroplasto “prestado”
En el corazón de la fotosíntesis se encuentra el Fotosistema I, un complejo proteína–pigmento que convierte la energía lumínica en el flujo de electrones necesario para sintetizar azúcares. En la mayoría de plantas y algas verdes, esta unidad central está rodeada por una media luna de proteínas antena que recogen la luz. Euglena gracilis es diferente: su cloroplasto se originó por la fagocitosis de un alga verde ya fotosintética, proporcionándole lo que los biólogos denominan un plastidio secundario. Ese origen poco habitual dejó en Euglena pigmentos y componentes proteicos distintivos, y hasta ahora no se conocía la disposición exacta de su supercomplejo del Fotosistema I. Utilizando crio-microscopía electrónica de alta resolución, los autores resolvieron la estructura tridimensional de este ensamblaje hasta 2,06 angstroms, revelando cómo Euglena ha reconstruido su motor lumínico.
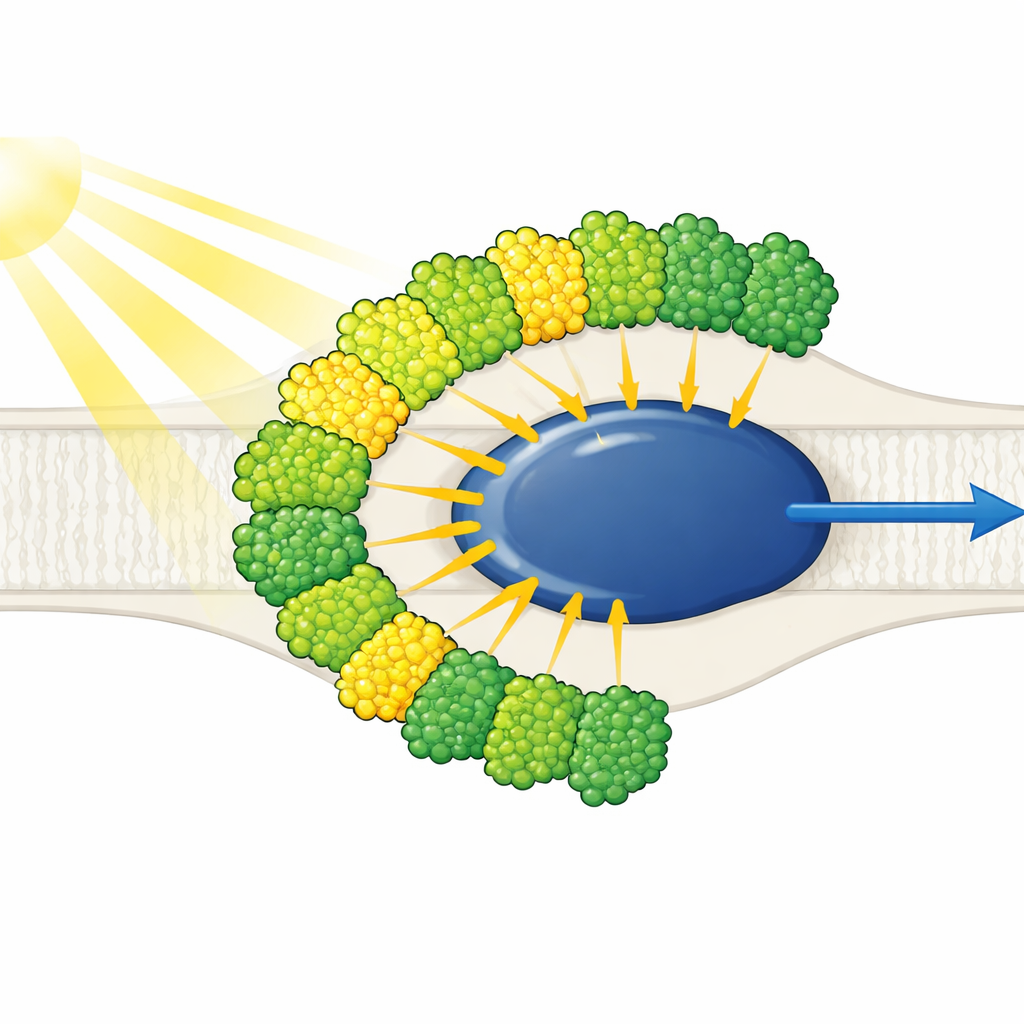
Antenas de luz todas en un mismo lado
La estructura muestra un núcleo del Fotosistema I reducido rodeado por diez antenas de captación de luz: nueve de un tipo (LHCI) y una normalmente asociada al Fotosistema II (LHCII). De forma llamativa, las diez antenas se ubican en un solo lado del núcleo, en dos cinturones concéntricos, en lugar de formar la típica media luna vegetal en el lado opuesto. Cada antena exterior se empareja cara a cara con una vecina interior, creando cinco dímeros apilados que a su vez se ensamblan en dos tetrámeros. Estas antenas están construidas a partir de poliproteínas que se cortan en unidades individuales, y motivos de secuencia cortos y específicos actúan como Velcro molecular que mantiene unidos los dímeros. Una antena interior, denominada LHC-3, emerge como un eje central, enlazando dímeros y ayudando a fijar todo el cinturón al núcleo.
Un nuevo lazo proteico que sustituye partes perdidas
En comparación con los sistemas vegetales familiares, el núcleo del Fotosistema I de Euglena carece de varias pequeñas proteínas ancla que normalmente ayudan a posicionar las antenas y permiten la reorganización dinámica ante cambios de luz. En su lugar, una de las subunidades del núcleo que conserva, PsaD, porta un lazo extra de alrededor de 40 aminoácidos que solo se encuentra en Euglena. Esta inserción flexible alcanza el espacio donde dichas proteínas ausentes se situarían normalmente y forma contactos estabilizadores directos con LHC-3 y sus clorofilas. Los autores proponen que este lazo único evolucionó junto con la pérdida de las subunidades ausentes, asumiendo su papel en sujetar las antenas al núcleo y condicionando dónde y cómo la energía lumínica entrante accede a la máquina.
Pigmentos a medida y autopistas de energía
Más allá de la arquitectura proteica, la posición de las moléculas de clorofila determina cómo fluye la energía lumínica. El complejo de Euglena contiene más de 200 pigmentos, incluidas clorofilas a y b y carotenoides especiales que difieren de los de las plantas terrestres. Dos clorofilas, etiquetadas a1020 y a1021 por analogía con sitios conocidos, son únicas de Euglena y aparecen solo en antenas interiores. Se sitúan en puntos de unión donde confluyen tres antenas, formando racimos de pigmentos que puentean físicamente unidades vecinas. Estos racimos, junto con pares de clorofilas “rojas” estrechamente espaciadas que absorben luz de energía ligeramente menor, crean múltiples rutas para que la excitación viaje desde el cinturón exterior al interior y luego al núcleo. Cálculos basados en las distancias entre centros de pigmentos muestran que la energía recogida en las antenas exteriores se canaliza preferentemente hacia unas pocas antenas de entrada, especialmente LHC-3 y su contraparte LHC-9, antes de ser transferida al centro de reacción donde tiene lugar la transferencia de electrones.
Enlaces flexibles para adaptarse a la luz cambiante
No todas las antenas están ancladas con la misma firmeza. La única unidad LHCII y una LHCI exterior muestran ocupancia parcial en los datos de crio-EM, lo que sugiere que pueden acoplarse y desacoplarse más fácilmente que las demás. Esta labilidad apunta a una característica reguladora: al añadir o quitar estas antenas, Euglena podría ajustar la cantidad de luz que alcanza el Fotosistema I y equilibrar la energía compartida con el Fotosistema II, evocando transiciones de estado conocidas en plantas pero usando una solución estructural diferente. Junto con sus pigmentos únicos y su disposición asimétrica de antenas, el diseño de Euglena parece optimizado para una captura de energía robusta pero ajustable.
Qué significa esto para la vida impulsada por el sol
Para un público general, el mensaje clave es que la evolución tiene muchas maneras de resolver el problema de convertir la luz solar en energía utilizable. Euglena gracilis, al haber adquirido su cloroplasto “de segunda mano”, no se limitó a copiar el esquema estándar vegetal. En su lugar, redujo el núcleo, reubicó todas las antenas a un lado, inventó un nuevo lazo proteico para anclarlas y desplegó racimos de clorofilas y pigmentos distintivos para encauzar la energía de forma eficiente hacia su centro de reacción. Esta solución propia de la línea evolutiva muestra cómo los plastidios secundarios pueden remodelar el aparato fotosintético mientras preservan —e incluso mejoran— la tarea básica de captar luz para la vida.
Cita: Huang, G., Dong, S., Li, L. et al. The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis. Nat Commun 17, 3429 (2026). https://doi.org/10.1038/s41467-026-70183-y
Palabras clave: Euglena gracilis, sistema fotosintético I, complejos antena de captación de luz, transferencia de energía por excitación, crio-microscopía electrónica