Clear Sky Science · nl
De architectuur en energietransferroutes van PSI–LHCI–LHCII in de phototrofe flagellaat Euglena gracilis
Hoe een piepkleine zwemmer zonlicht in energie omzet
De eencellige alg Euglena gracilis is een microscopische zwemmer die goed gedijt in vijvers en plassen. Net als planten gebruikt zij zonlicht om het leven van energie te voorzien, maar ze doet dat met een fotosynthetisch apparaat dat een ongewone evolutionaire weg heeft afgelegd. Deze studie onthult dat apparaat tot atomair detail en laat zien hoe Euglena zijn lichtopvangsysteem heeft herbedraad om energie efficiënt en flexibel te vangen, nadat het miljoenen jaren geleden zijn chloroplast van een andere alg verwierf.
Een speciaal lichtmotor in een geleend chloroplast
In het hart van de fotosynthese ligt Fotosysteem I, een eiwit–pigmentcomplex dat lichtenergie omzet in een elektronenstroom die nodig is om suikers te bouwen. Bij de meeste planten en groene algen wordt deze kern omgeven door een halvemaan van antenne-eiwitten die licht verzamelen. Euglena gracilis verschilt: haar chloroplast ontstond door het opnemen van een al fotosynthetische groene alg, waardoor zij wat biologen een secundaire plastide noemen. Die ongewone oorsprong liet bij Euglena karakteristieke pigmenten en eiwitcomponenten achter, en tot nu toe was de precieze opstelling van haar Fotosysteem I-supercomplex onbekend. Met hoge-resolutie kryo-elektronenmicroscopie brachten de auteurs de driedimensionale structuur in kaart tot 2,06 ångström en toonden ze hoe Euglena zijn lichtmotor heeft herbouwd.
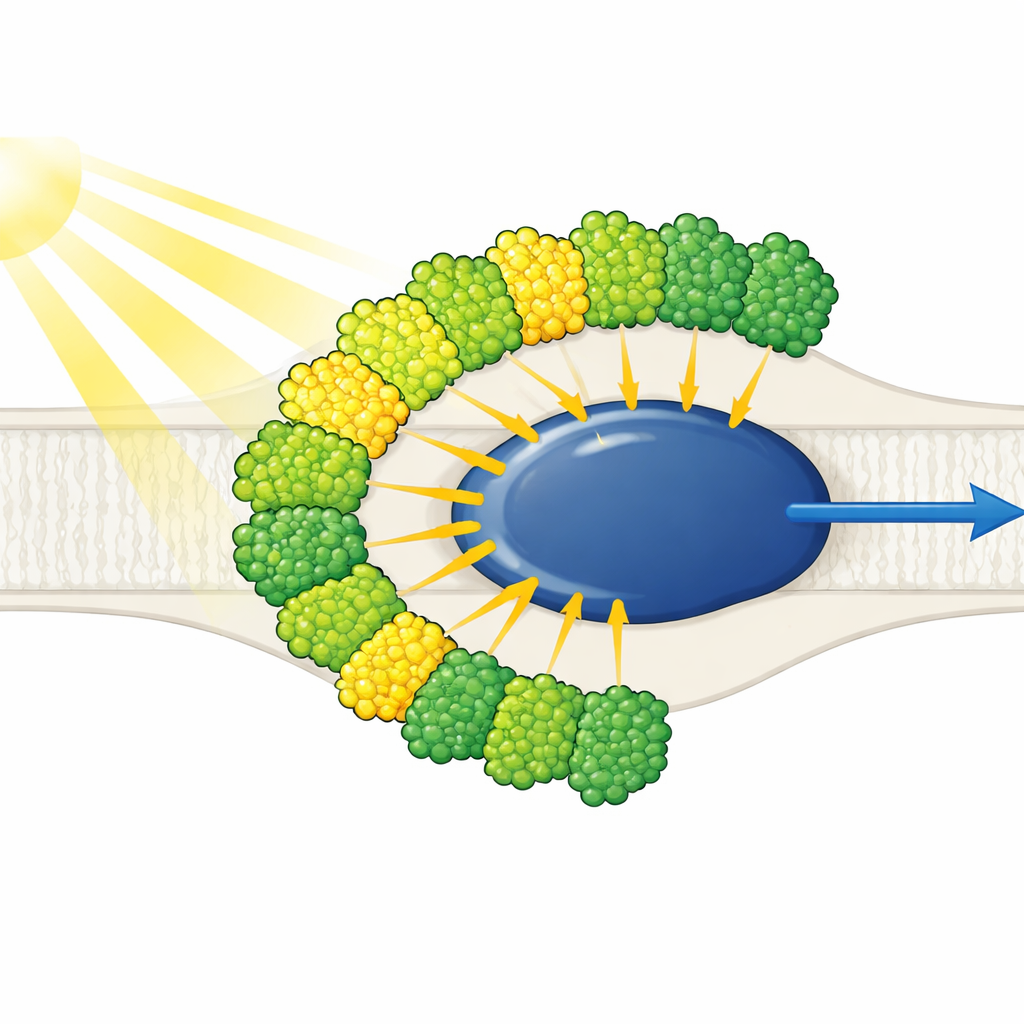
Lichtantennes aan één kant
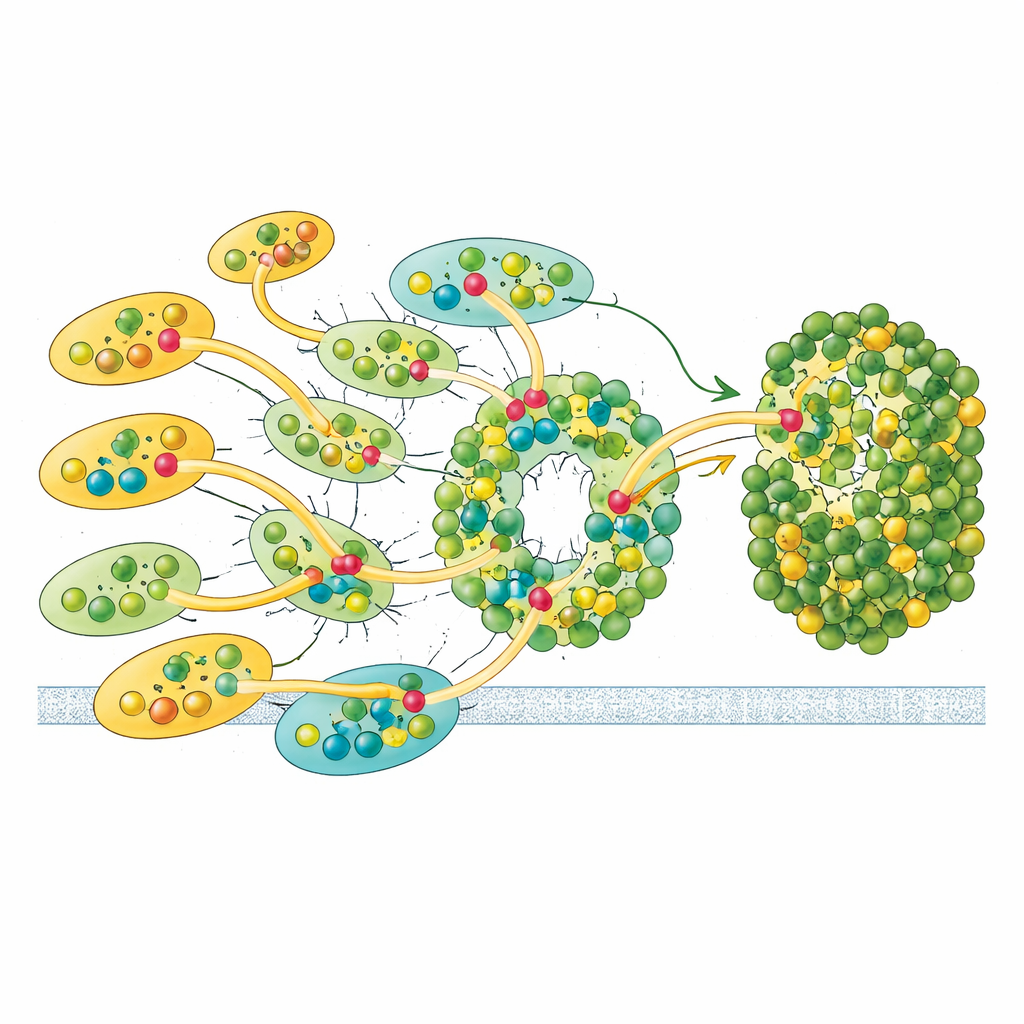
De structuur toont een afgeslankte Fotosysteem I-kern omringd door tien lichtopvangantennes—negen van het ene type (LHCI) en één die gewoonlijk met Fotosysteem II geassocieerd wordt (LHCII). Opmerkelijk is dat alle tien antennes aan slechts één kant van de kern zitten, in twee concentrische rijen, in plaats van de typische plantachtige halvemaan aan de tegenoverliggende zijde. Iedere buitenste antenne vormt een face-to-facepaar met een innerlijke buur, waardoor vijf gestapelde dimeren ontstaan die zich op hun beurt assembleren tot twee tetrameren. Deze antennes zijn opgebouwd uit polyproteïnen die worden opgesplitst in individuele eenheden, en specifieke korte sequentiemotieven werken als moleculaire klittenband die de dimeren bij elkaar houden. Eén innerlijke antenne, LHC-3 genoemd, blijkt een centraal knooppunt te zijn dat dimeren linkt en helpt de hele rij aan de kern te verankeren.
Een nieuwe proteïnelus ter vervanging van verloren onderdelen
Vergeleken met bekende plantsystemen ontbreekt in Euglena’s Fotosysteem I-kern een aantal kleine ankerproteïnen die normaal gesproken antennes positioneren en dynamische reorganisatie onder veranderend licht mogelijk maken. In plaats daarvan draagt een van de overgebleven kernsubunits, PsaD, een extra lus van ongeveer 40 aminozuren die alleen in Euglena voorkomt. Deze flexibele insertie reikt in de ruimte waar de ontbrekende eiwitten normaal zouden zitten en vormt directe stabiliserende contacten met LHC-3 en diens chlorofylen. De auteurs stellen voor dat deze unieke lus samen met het verlies van de afwezige subunits is geëvolueerd en hun rol heeft overgenomen bij het bevestigen van antennes aan de kern en het bepalen waar en hoe binnenkomende lichtenergie het complex binnenkomt.
Aangepaste pigmenten en energiewegen
Naast de eiwitarchitectuur bepaalt de positionering van chlorofylmoleculen hoe lichtenergie stroomt. Het Euglena-complex bevat meer dan 200 pigmenten, waaronder chlorofylen a en b en speciale carotenoïden die verschillen van die in landplanten. Twee chlorofylen, analoog gelabeld als a1020 en a1021, zijn uniek voor Euglena en komen alleen in innerlijke antennes voor. Ze zitten op kruispunten waar drie antennes samenkomen en vormen pigmentclusters die fysiek naburige eenheden overbruggen. Deze clusters, samen met nauw geplaatste “rode” chlorofylparen die iets lagere-energie licht absorberen, creëren meerdere routes waarlangs excitatie kan reizen van de buitenste rij naar de binnenste rij en vervolgens naar de kern. Berekeningen gebaseerd op afstanden tussen pigmentcentra laten zien dat energie die in buitenste antennes wordt verzameld, bij voorkeur wordt geleid naar een paar poortantennes, vooral LHC-3 en zijn tegenhanger LHC-9, voordat ze worden doorgegeven aan het reactiemembraan waar elektronenoverdracht plaatsvindt.
Flexibele verbindingen voor aanpassing aan veranderend licht
Niet alle antennes zijn even vast bevestigd. De enige LHCII-eenheid en één buitenste LHCI tonen slechts gedeeltelijke bezetting in de kryo-EM-gegevens, wat suggereert dat ze makkelijker kunnen aan- en losmaken dan de andere. Deze labiele binding wijst op een regulerend mechanisme: door antennes toe te voegen of te verwijderen kan Euglena mogelijk afstellen hoeveel licht Fotosysteem I bereikt en de balans van energie met Fotosysteem II aanpassen, een echo van staatstransities die bij planten bekend zijn, maar bereikt via een andere structurele oplossing. Samen met zijn unieke pigmenten en asymmetrische antenne-opstelling lijkt Euglena’s ontwerp geoptimaliseerd voor robuuste maar aanpasbare energieopname.
Wat dit betekent voor leven op zonne-energie
Voor niet-specialisten is de kernboodschap dat evolutie vele manieren kent om het probleem van het omzetten van zonlicht in bruikbare energie op te lossen. Euglena gracilis, die haar chloroplast “tweedehands” verwierf, kopieerde niet eenvoudig het standaard plantontwerp. In plaats daarvan heeft ze de kern afgeslankt, alle antennes aan één kant gepositioneerd, een nieuwe proteïnelus uitgevonden om ze te verankeren, en karakteristieke chlorofylclusters en pigmenten ingezet om energie efficiënt naar haar reactievergadering te sturen. Deze lijnspecifieke oplossing laat zien hoe secundaire plastiden het fotosynthetische apparaat kunnen herschikken terwijl ze de fundamentele taak van lichtoogst voor het leven behouden—en zelfs verbeteren.
Bronvermelding: Huang, G., Dong, S., Li, L. et al. The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis. Nat Commun 17, 3429 (2026). https://doi.org/10.1038/s41467-026-70183-y
Trefwoorden: Euglena gracilis, fotosysteem I, lichtopvangcomplexen, excitatiesenergietransfer, kryo-elektronenmicroscopie