Clear Sky Science · fr
L’architecture et les voies de transfert d’énergie du PSI–LHCI–LHCII chez le flagellé phototrophe Euglena gracilis
Comment un minuscule nageur transforme la lumière du soleil en énergie
L’algue unicellulaire Euglena gracilis est un nageur microscopique qui prospère dans les étangs et les flaques. Comme les plantes, elle utilise la lumière du soleil pour alimenter la vie, mais elle le fait avec une machinerie photosynthétique qui a suivi une voie évolutive inhabituelle. Cette étude révèle cette machinerie au niveau atomique, montrant comment Euglena a reconfiguré son système de collecte de lumière pour capter l’énergie de façon efficace et flexible après avoir acquis son chloroplaste d’une autre algue il y a des millions d’années.
Un moteur lumineux particulier dans un chloroplaste emprunté
Au cœur de la photosynthèse se trouve le photosystème I, un complexe protéine–pigment qui convertit l’énergie lumineuse en flux d’électrons nécessaires à la synthèse des sucres. Chez la plupart des plantes et des algues vertes, cette unité centrale est entourée d’un croissant de protéines antennes qui collectent la lumière. Euglena gracilis est différente : son chloroplaste provient de l’engloutissement d’une algue verte déjà photosynthétique, lui conférant ce que les biologistes appellent un plastide secondaire. Cette origine inhabituelle a laissé à Euglena des pigments et des composants protéiques distinctifs, et jusqu’à présent, la configuration exacte de son supercomplexe du photosystème I était inconnue. Grâce à la cryomicroscopie électronique à haute résolution, les auteurs ont résolu la structure tridimensionnelle de cet assemblage jusqu’à 2,06 angströms, révélant comment Euglena a reconstruit son moteur lumineux.
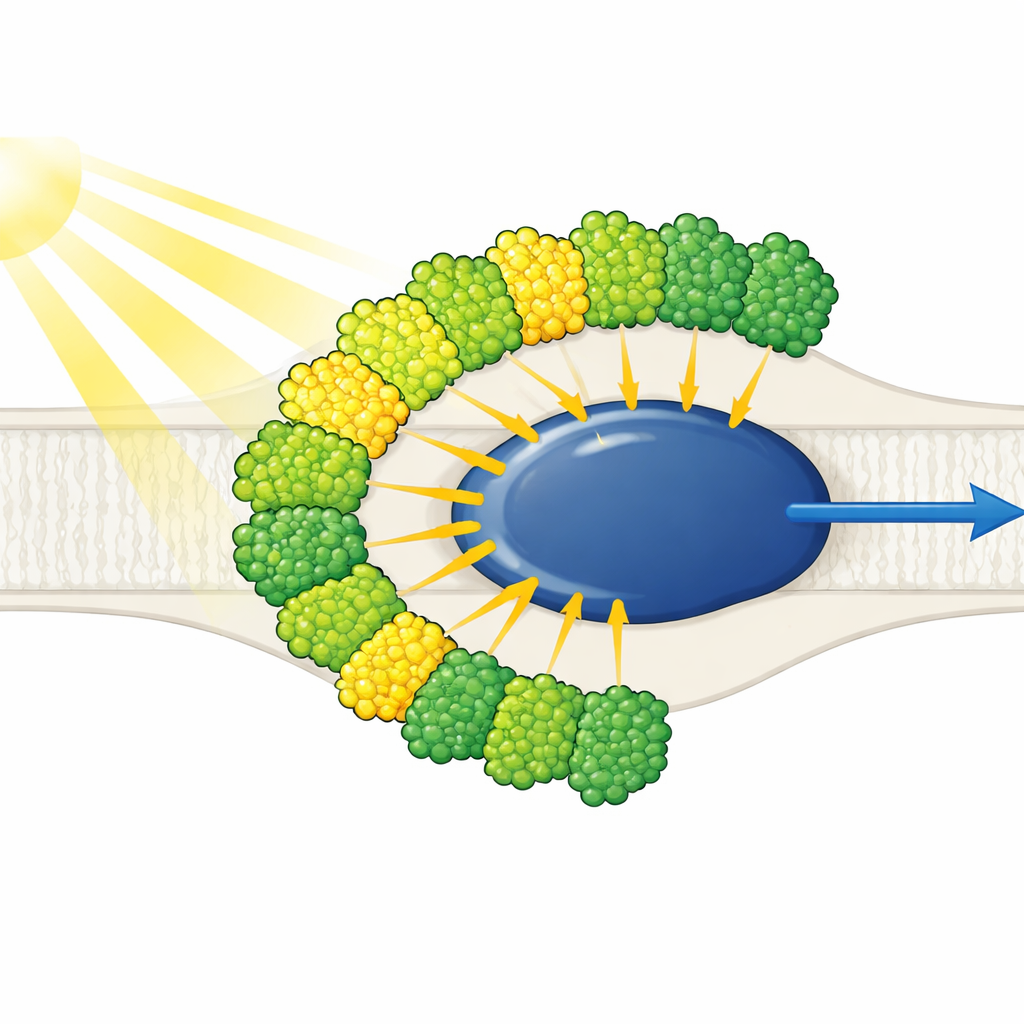
Des antennes lumineuses toutes d’un même côté
La structure montre un cœur de photosystème I épuré entouré de dix antennes collectrices de lumière — neuf d’un type (LHCI) et une généralement associée au photosystème II (LHCII). Fait remarquable, les dix antennes se trouvent du même côté du cœur, disposées en deux ceintures concentriques, plutôt que de former le croissant typique semblable à celui des plantes de l’autre côté. Chaque antenne externe fait face à une voisine interne, créant cinq dimères empilés qui s’assemblent à leur tour en deux tétramères. Ces antennes sont constituées de polyprotéines scindées en unités individuelles, et des motifs de séquence courts spécifiques fonctionnent comme du Velcro moléculaire maintenant les dimères ensemble. Une antenne interne, appelée LHC-3, émerge comme un noyau central, reliant les dimères et aidant à fixer l’ensemble de la ceinture au cœur.
Une nouvelle boucle protéique remplaçant des éléments perdus
Comparé aux systèmes végétaux familiers, le cœur du photosystème I d’Euglena manque de plusieurs petites protéines d’ancrage qui aident généralement à positionner les antennes et permettent la réorganisation dynamique sous un éclairage changeant. À la place, l’une de ses sous-unités centrales restantes, PsaD, porte une boucle supplémentaire d’environ 40 acides aminés trouvée uniquement chez Euglena. Cette insertion flexible s’étend dans l’espace où les protéines absentes siégeraient normalement et forme des contacts stabilisants directs avec LHC-3 et ses chlorophylles. Les auteurs proposent que cette boucle unique ait évolué parallèlement à la perte des sous-unités manquantes, prenant en charge leur rôle pour fixer les antennes au cœur et déterminer où et comment l’énergie lumineuse entrante pénètre dans la machinerie.
Pigments sur mesure et autoroutes énergétiques
Au-delà de l’architecture protéique, la manière dont les molécules de chlorophylle sont positionnées détermine le flux d’énergie lumineuse. Le complexe d’Euglena contient plus de 200 pigments, y compris les chlorophylles a et b et des caroténoïdes spéciaux différents de ceux des plantes terrestres. Deux chlorophylles, nommées a1020 et a1021 par analogie à des sites connus, sont uniques à Euglena et apparaissent uniquement dans les antennes internes. Elles se situent à des jonctions où trois antennes se rencontrent, formant des amas de pigments qui relient physiquement les unités voisines. Ces amas, avec des paires de chlorophylles « rouges » très rapprochées absorbant une lumière légèrement moins énergétique, créent plusieurs voies par lesquelles l’excitation peut voyager de la ceinture externe vers la ceinture interne puis vers le cœur. Des calculs basés sur les distances entre centres de pigments montrent que l’énergie collectée dans les antennes externes est acheminée préférentiellement vers quelques antennes porteuses, en particulier LHC-3 et son homologue LHC-9, avant d’être transférée au centre de réaction où se produit le transfert d’électrons.
Liens flexibles pour s’adapter à la lumière changeante
Toutes les antennes ne sont pas fixées avec la même rigidité. L’unité unique LHCII et une antenne externe LHCI présentent une occupation partielle dans les données cryo-EM, suggérant qu’elles peuvent se fixer et se détacher plus facilement que les autres. Cette labilité suggère une fonction régulatrice : en ajoutant ou retirant ces antennes, Euglena peut moduler la quantité de lumière atteignant le photosystème I et ajuster l’équilibre d’énergie partagé avec le photosystème II, rappelant les transitions d’état connues chez les plantes mais par une solution structurelle différente. Avec ses pigments uniques et sa disposition asymétrique des antennes, le design d’Euglena semble optimisé pour une capture d’énergie robuste mais modulable.
Ce que cela signifie pour la vie propulsée par le soleil
Pour un non-spécialiste, le message principal est que l’évolution propose de nombreuses solutions au problème de convertir la lumière du soleil en énergie utilisable. Euglena gracilis, ayant acquis son chloroplaste « d’occasion », n’a pas simplement copié le plan type des plantes. Elle a au contraire simplifié le cœur, repositionné toutes les antennes d’un même côté, inventé une nouvelle boucle protéique pour les ancrer, et déployé des clusters de chlorophylles et des pigments distinctifs pour diriger l’énergie efficacement vers son centre de réaction. Cette solution spécifique à une lignée montre comment les plastides secondaires peuvent remodeler l’appareil photosynthétique tout en préservant — et même en améliorant — la fonction essentielle de récolter la lumière pour la vie.
Citation: Huang, G., Dong, S., Li, L. et al. The architecture and energy transfer pathways of PSI–LHCI–LHCII in the phototrophic flagellate Euglena gracilis. Nat Commun 17, 3429 (2026). https://doi.org/10.1038/s41467-026-70183-y
Mots-clés: Euglena gracilis, photosystème I, complexes collecteurs de lumière, transfert d’énergie d’excitation, cryomicroscopie électronique