Clear Sky Science · zh
UBA6 的冷冻电镜结构揭示了 E1–E2 特异性与双重 FAT10/泛素硫酯转移的机制
细胞如何决定为哪些蛋白打标签
在每个细胞内部,一套隐秘的标签系统决定哪些蛋白被送去回收、哪些被重塑、以及哪些参与免疫反应。两种微小的标签——泛素和 FAT10——参与这些决策,它们必须以极高的精确度加载到载体酶上。本文探讨了一个关键的加载酶 UBA6 如何选择性地与特定的合作酶配合并同时处理泛素和 FAT10,揭示了维持细胞蛋白控制系统准确且可调的设计原理。
两条并行的标记通道
细胞使用分子标签来标记蛋白,以决定其不同命运,从降解到活性或定位的变化。泛素是最为人所知的标签,参与多种细胞过程,而 FAT10 则更为专一,与免疫反应和炎症密切相关。两种标签都是通过包括 E1、E2 和 E3 酶的三级接力附着到靶蛋白上。E1 酶执行第一步消耗能量的激活步骤,将标签活化并传递给 E2,随后 E2 与 E3 合作将标签连接到靶蛋白上。人类在泛素通路上具有两个 E1 酶:负责经典泛素通路的 UBA1,以及不仅激活泛素还可激活 FAT10 的 UBA6,后者在日常蛋白控制与免疫调控的蛋白处置之间架起了一座桥梁。
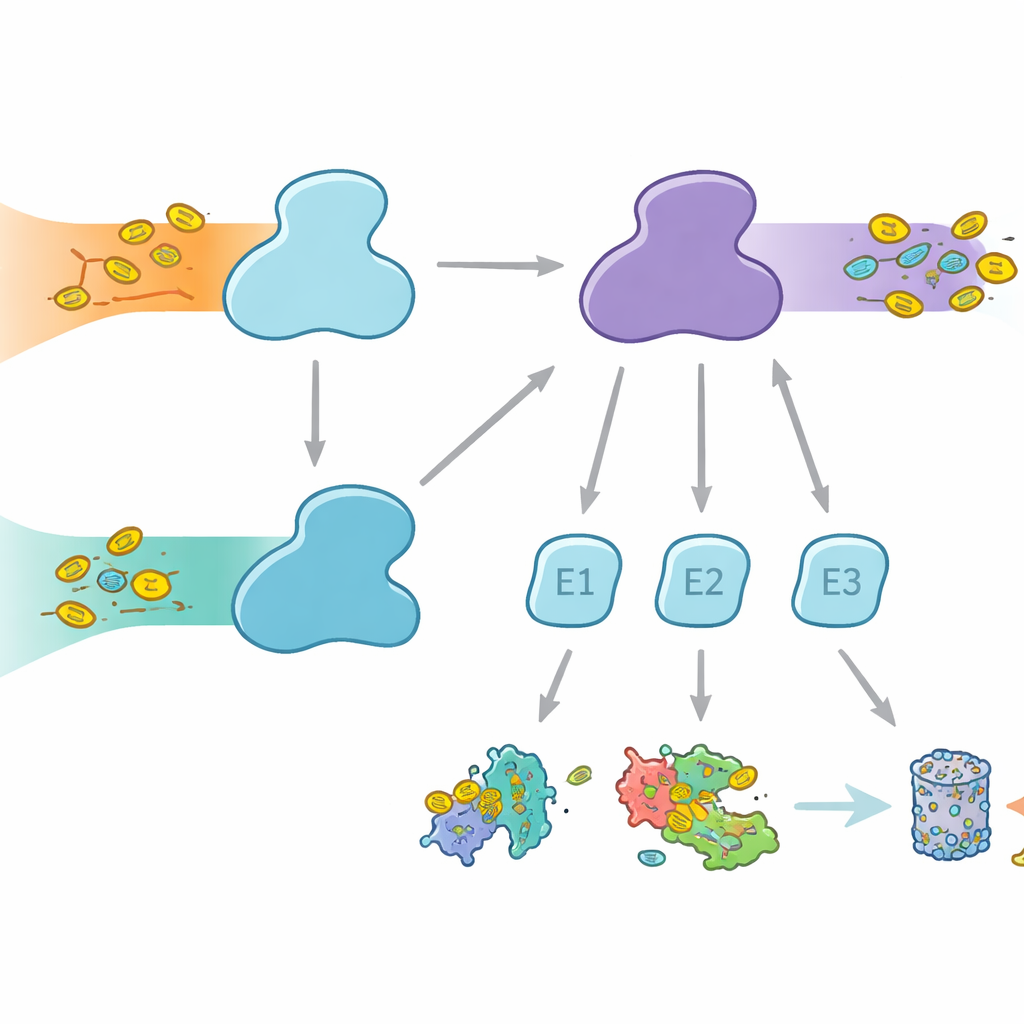
为何选择合作伙伴很重要
尽管 UBA1 与 UBA6 在总体上看起来相似,但它们与大体不同的 E2 合作酶集合配对,形成了标记系统的并行分支。有些 E2 可以同时与两者配合,但也有些是专属伙伴。这种选择性至关重要:若形成了错误的组合,免疫相关的 FAT10 信号可能泄入常规的泛素通路,或反之,从而扰乱细胞决策。此前尚不清楚 UBA6 如何识别其偏好的 E2,特别是一组被称为 IV 类的专门 E2,而 UBA1 在很大程度上忽视它们。作者们利用高分辨率冷冻电子显微镜和高灵敏度的生化测定,试图捕捉 UBA6 与其合作伙伴在作用中的形态与动力学。
双手抓握的识别策略
结构显示 UBA6 使用一种“二手握持”方式识别其关键伙伴 E2,即 UBE2Z。一只“手”是位于 UBA6 末端、为 E2 提供停靠位的结构域,另一只“手”是更靠近化学反应位点的催化区域。与主要依赖停靠区选择伙伴的 UBA1 不同,UBA6 将两者协同使用,以将 UBE2Z 优雅地包裹在能精确对齐进行标签转移所需的两个反应位点之间。UBE2Z 上一些其他 E2 所没有的额外环状片段恰好插入 UBA6 扩大的沟槽,使得结合既紧密又具有柔性。如果停靠表面或催化裂缝中的任一部分发生改变,UBA6 就无法有效装载 UBE2Z,表明两只“手”对正确的伙伴选择都是必须的。
一个小型辅因子与一个灵活的开关
一个令人意外的发现是,一种称为六磷酸肌醇的小分子嵌入在靠近 UBA6 催化裂缝的口袋中。该辅因子有助于将沟槽保持在一种欢迎 IV 类 E2(包括 UBE2Z 和 BIRC6)不同寻常环段的开放构象,同时使同一区域对 UBA1 偏好的 E2 变得不那么相容。对一个中间态的放大结构显示了当 UBA6 与 UBE2Z 同时与两个泛素或两个 FAT10 分子结合时标签如何被移交:UBE2Z 中一个可移动的环像门一样摆开,暴露其反应性的半胱氨酸以便转移标签。接触界面能够适应不同化学性质:与 FAT10 形成时更具极性、基于电荷的界面,而与泛素形成时则更偏疏水、类油性的界面,这使得同一 E2 能够处理两种化学上不同的标签。
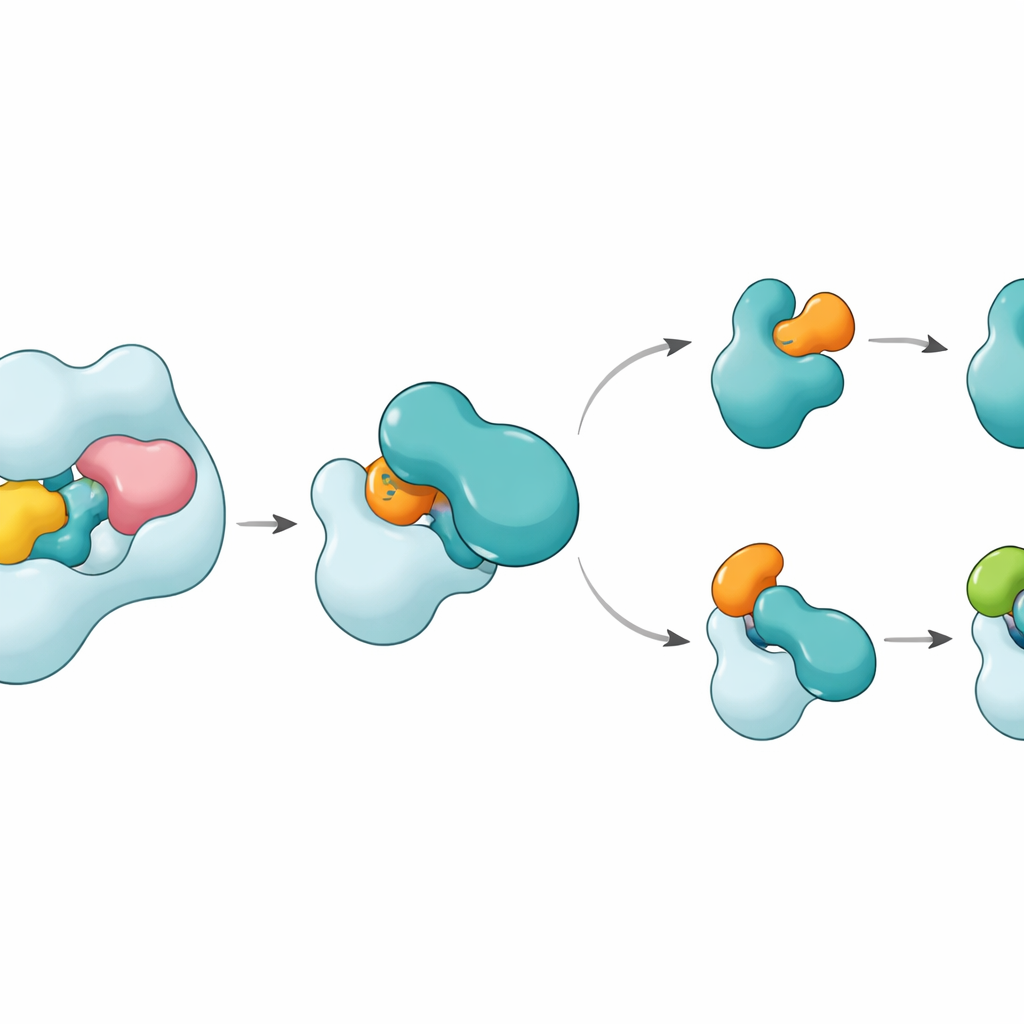
这如何让细胞的标签系统保持正轨
简而言之,这项工作表明 UBA6 已进化出一种特化的、由两部分组成的识别摇篮,并由小型辅因子加固,使其能够与一组精选的 E2 伙伴紧密协作并同时支持泛素与 FAT10 的标记。相比之下,UBA1 则采用更传统的单一区域策略并与另一套 E2 配对。这些不同的设计方案在细胞内形成了两条并行但相互绝缘的标记分支:一条处理日常的蛋白质控制,另一条将蛋白处置与免疫信号连接。理解这种架构逻辑不仅阐明了细胞如何在其蛋白网络中维持秩序,也可能为未来在癌症、免疫及其他疾病中靶向标记系统特定分支的努力提供参考。
引用: Nayak, D., Jia, L., dos Santos Bury, P. et al. Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer. Nat Commun 17, 3302 (2026). https://doi.org/10.1038/s41467-026-69882-3
关键词: 泛素, FAT10, UBA6, 蛋白质降解, 冷冻电镜