Clear Sky Science · nl
Cryo-EM-structuren van UBA6 onthullen mechanismen van E1–E2-specificiteit en dubbele FAT10/ubiquitine-thioesteroverdracht
Hoe cellen beslissen welke eiwitten worden gemarkeerd
Binnen elke cel bepaalt een onzichtbaar labelingssysteem welke eiwitten naar recycling worden gestuurd, welke worden heringericht en welke helpen bij het opwekken van een immuunrespons. Twee kleine labels, ubiquitine en FAT10, helpen deze beslissingen te nemen en moeten met grote precisie op draagende enzymen worden geladen. Dit artikel onderzoekt hoe één belangrijke laadenzym, genaamd UBA6, selectief samenwerkt met bepaalde partner‑enzymen en zowel ubiquitine als FAT10 verwerkt, en onthult ontwerprichtlijnen die het eiwitbeheersysteem van de cel nauwkeurig en aanpasbaar houden.
Twee parallelle labelingssnelwegen
Cellen gebruiken moleculaire labels om eiwitten voor verschillende bestemmingen te markeren, van afbraak tot veranderingen in activiteit of locatie. Ubiquitine is het bekendste label en wordt in veel cellulaire processen gebruikt, terwijl FAT10 specialistischer is en sterk verbonden is met immuunreacties en ontsteking. Beide labels worden bevestigd via een drie‑stappenrelais met E1-, E2‑ en E3‑enzymen. De E1‑enzymen voeren de eerste, energieverslindende stap uit: ze activeren het label en geven het door aan E2’s, die vervolgens met E3’s samenwerken om het label aan doelwit‑eiwitten te koppelen. Mensen zijn bijzonder doordat ze twee E1‑enzymen voor ubiquitine hebben: UBA1, die het klassieke ubiquitinepad bedient, en UBA6, die niet alleen ubiquitine activeert maar ook FAT10, en daarmee een brug vormt tussen alledaagse eiwitcontrole en immuungecontrole van eiwitafvoer.
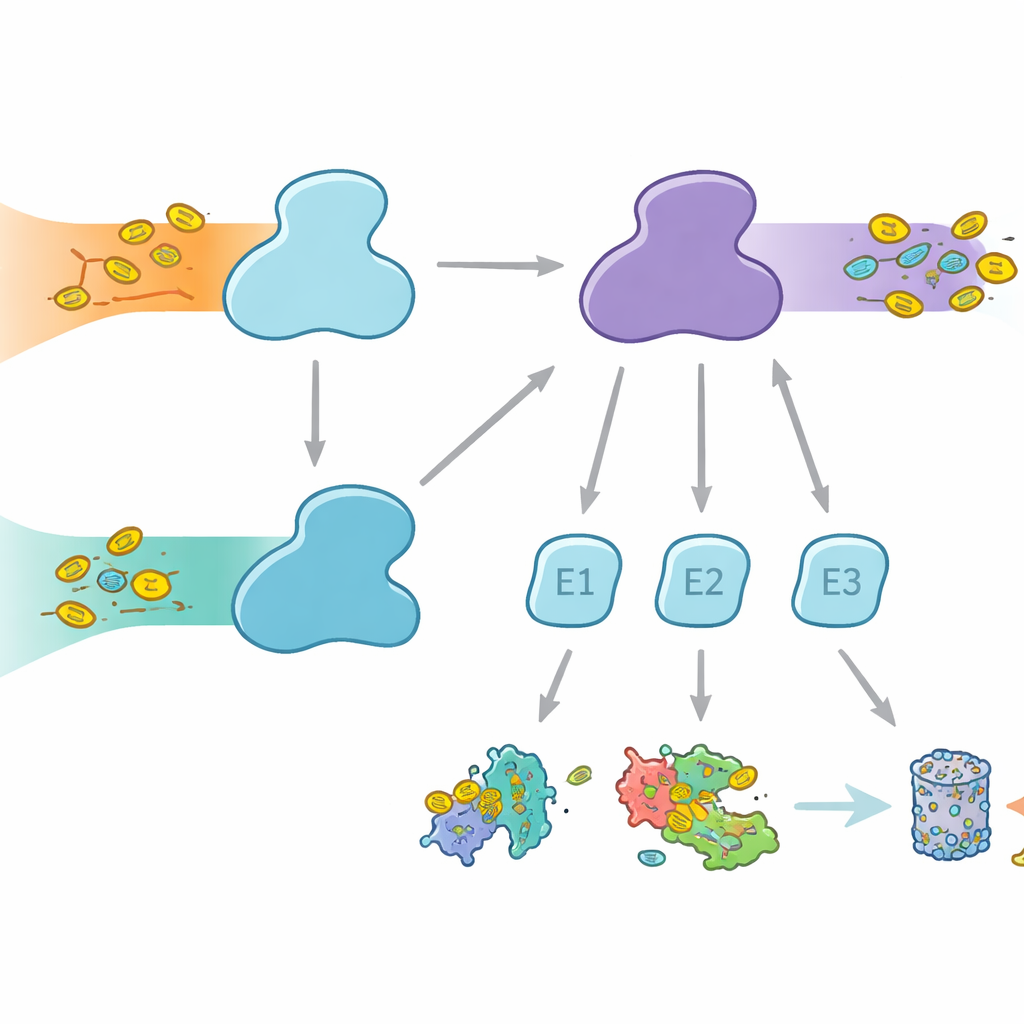
Waarom partnerkeuze ertoe doet
Hoewel UBA1 en UBA6 in grote lijnen op elkaar lijken, werken ze met grotendeels verschillende sets E2‑partners, waardoor parallelle takken van het labelingssysteem ontstaan. Sommige E2’s kunnen met beide E1’s werken, maar andere zijn exclusieve partners. Deze selectiviteit is cruciaal: als de verkeerde combinaties zouden ontstaan, zouden immuun‑gekoppelde FAT10‑signalen kunnen weglekken naar gewone ubiquitinepaden of omgekeerd, wat cellulaire beslissingen zou verstoren. Tot nu toe was onduidelijk hoe UBA6 zijn voorkeurs‑E2’s herkent, vooral een gespecialiseerde groep die bekendstaat als klasse IV E2’s, terwijl UBA1 deze grotendeels negeert. De auteurs probeerden UBA6 en zijn partners tijdens het werk vast te leggen met hoge‑resolutie cryo‑elektronenmicroscopie en biochemische experimenten die de labeloverdracht zeer gevoelig meten.
Een herkenningsstrategie met twee grepen
De structuren tonen aan dat UBA6 zijn belangrijkste partner E2, genaamd UBE2Z, herkent met een "tweehandige" greep. De ene hand is een domein aan het uiteinde van UBA6 dat als aanlegplaats voor E2’s dient, terwijl de andere hand een katalytisch gebied is dat dichter bij de reactieweg ligt. In tegenstelling tot UBA1, dat voornamelijk op het aanleggebied vertrouwt om partners te selecteren, gebruikt UBA6 beide regio’s samen om UBE2Z zodanig te omsluiten dat de twee reactieve sites voor labeloverdracht precies uitgelijnd zijn. Extra lussegmenten op UBE2Z, afwezig in veel andere E2’s, passen in een vergrote groef in UBA6, waardoor een strakke maar flexibele binding mogelijk is. Als ofwel het aanlegoppervlak ofwel de katalytische spleet wordt gewijzigd, slaagt UBA6 er niet in UBE2Z efficiënt te laden, wat aantoont dat beide "handen" nodig zijn voor correcte partnerkeuze.
Een kleine cofactor en een flexibele schakelaar
Een verrassing is dat een klein molecuul, inositolhexakisfosfaat, in een pocket nabij UBA6’s katalytische spleet past. Deze cofactor helpt de groef in een configuratie open te houden die de ongewone lussen van klasse IV E2’s, waaronder UBE2Z en BIRC6, verwelkomt, terwijl hetzelfde gebied minder compatibel wordt met E2’s die door UBA1 worden bevoordeeld. Ingekaderde structuren van een tussenstaat, waarin UBA6 en UBE2Z gelijktijdig gebonden zijn aan twee kopieën van ofwel ubiquitine ofwel FAT10, laten zien hoe het label wordt overgedragen. Een beweegbare lus in UBE2Z zwaait opzij als een poort en maakt zijn reactieve cysteïne bloot zodat het label kan worden overgedragen. Het contactoppervlak kan zich aanpassen: het vormt een meer polair, lading‑gebaseerd interfase met FAT10 en een meer hydrofoob, olieachtig interfase met ubiquitine, waardoor dezelfde E2 twee chemisch verschillende labels kan verwerken.
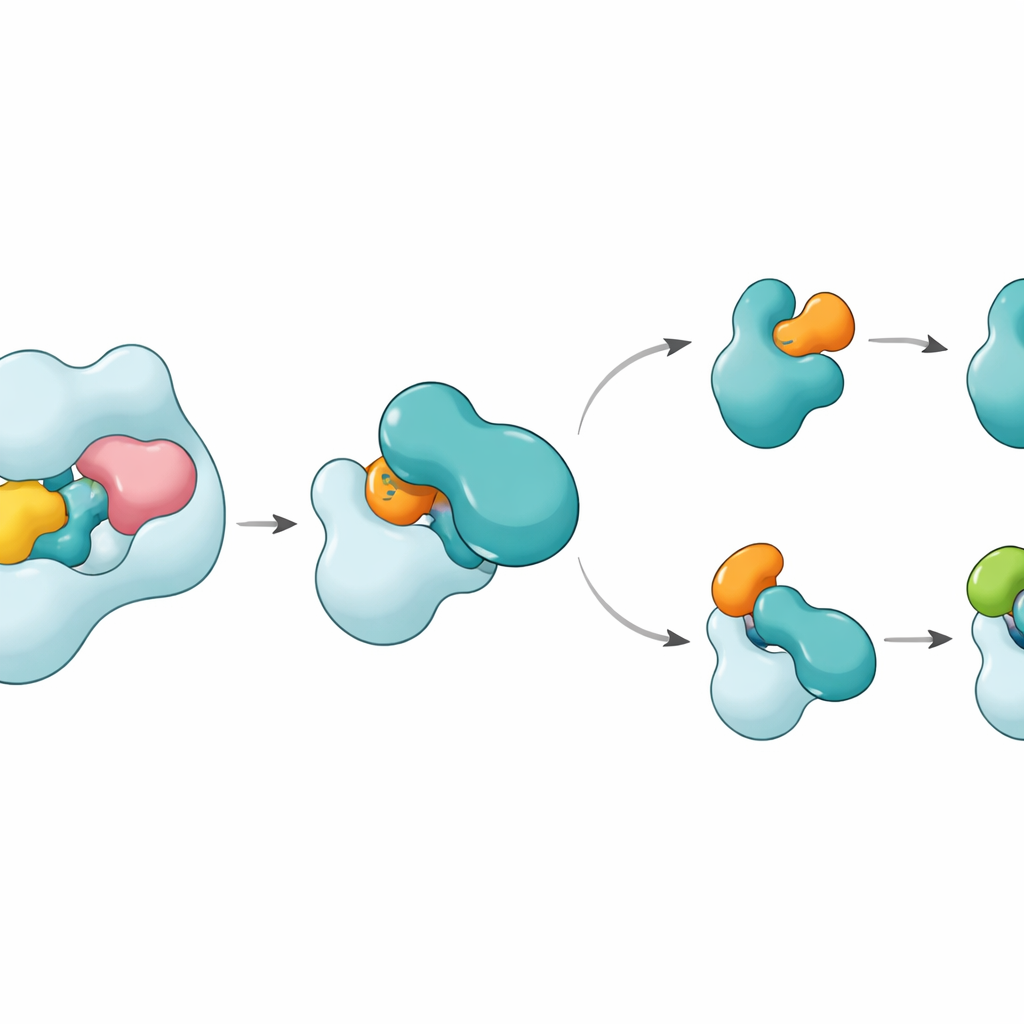
Hoe dit labelen in de cel op koers houdt
In eenvoudige bewoordingen laat dit werk zien dat UBA6 een gespecialiseerde, tweedelige herkenningskroes heeft ontwikkeld, versterkt door een kleine cofactor, die het in staat stelt nauw samen te werken met een selecte groep E2‑partners en zowel ubiquitine‑ als FAT10‑labeling te ondersteunen. UBA1 gebruikt daarentegen een meer traditionele, enkelregio‑strategie en werkt samen met een andere set E2’s. Deze verschillende ontwerpoplossingen creëren twee parallelle maar geïsoleerde labeltakken binnen de cel: één voor alledaagse eiwitcontrole en de andere die eiwitafvoer aan immuunsignalen koppelt. Inzicht in deze architectonische logica verduidelijkt niet alleen hoe cellen orde in hun eiwitnetwerken bewaren, maar kan ook toekomstige pogingen informeren om specifieke takken van het labelingssysteem te richten bij kanker, immuniteit en andere ziekten.
Bronvermelding: Nayak, D., Jia, L., dos Santos Bury, P. et al. Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer. Nat Commun 17, 3302 (2026). https://doi.org/10.1038/s41467-026-69882-3
Trefwoorden: ubiquitine, FAT10, UBA6, eiwitafbraak, cryo-EM