Clear Sky Science · sv
Cryo-EM-strukturer av UBA6 avslöjar mekanismer för E1–E2-specificitet och dubbel FAT10/ubiquitin-thioesteröverföring
Hur celler avgör vilka proteiner som märks
Inuti varje cell finns ett dolt märkningssystem som avgör vilka proteiner som skickas för återvinning, vilka som omformas och vilka som hjälper till att sätta igång ett immunsvar. Två små märkningar, ubiquitin och FAT10, bidrar till dessa beslut, och de måste laddas på bärarenzymer med utsökt precision. Denna artikel undersöker hur ett nyckelenzym för inladdning, kallat UBA6, selektivt samarbetar med vissa partnerenzym och hanterar både ubiquitin och FAT10, och avslöjar designprinciper som håller cellens proteinkontrollsystem både korrekt och anpassningsbart.
Två parallella märkningsvägar
Celler använder molekylära märkningar för att markera proteiner för olika öden, från nedbrytning till förändrad aktivitet eller lokalisering. Ubiquitin är den mest kända märkningen, använd i många cellulära processer, medan FAT10 är mer specialiserad och starkt kopplad till immunsvar och inflammation. Båda märkningarna fästs via ett trestegsrelä som involverar E1-, E2- och E3-enzymer. E1-enzymerna utför det första, energi‑krävande steget, aktiverar märkningen och överför den till E2:or, som sedan samarbetar med E3:or för att fästa märkningen på målproteiner. Människor är ovanliga genom att ha två E1-enzymer för ubiquitin: UBA1, som betjänar den klassiska ubiquitinvägen, och UBA6, som inte bara aktiverar ubiquitin utan också FAT10, och därigenom bildar en bro mellan vardaglig proteinstyrning och immunreglerad proteinbortskaffning.
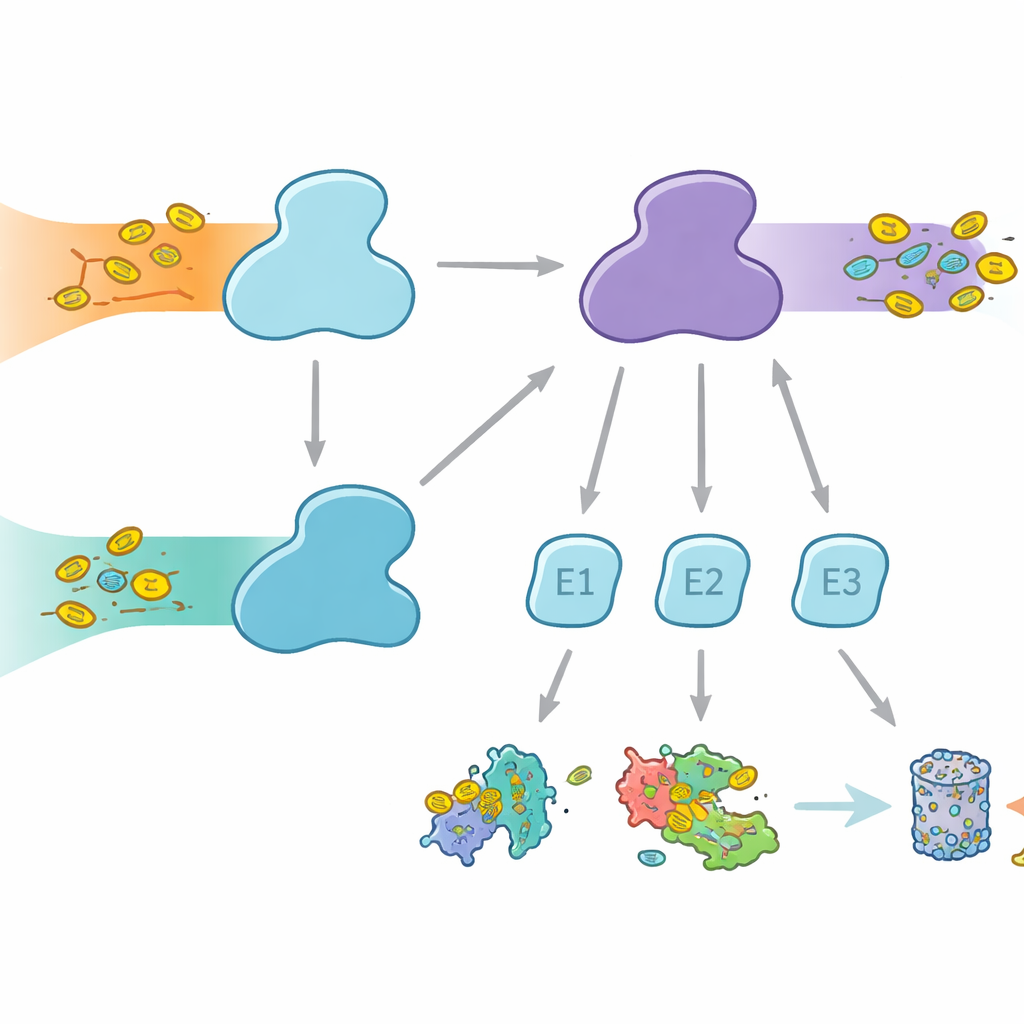
Varför partnerval spelar roll
Även om UBA1 och UBA6 ser i stora drag likartade ut, samarbetar de med i huvudsak olika uppsättningar av E2-partnerenzym, vilket skapar parallella grenar i märkningssystemet. Vissa E2:or kan arbeta med båda E1:orna, men andra är exklusiva partner. Denna selektivitet är avgörande: om fel kombinationer bildades skulle immunkopplade FAT10-signaler kunna läcka in i vanliga ubiquitinvägar, eller tvärtom, vilket skulle röra till cellens beslutsfattande. Hittills var det oklart hur UBA6 kunde känna igen sina föredragna E2:or, särskilt en specialiserad grupp känd som klass IV E2:or, medan UBA1 i stort ignorerar dem. Författarna satte upp att fånga UBA6 och dess partners in action med högupplöst kryo-elektronmikroskopi och biokemiska experiment som mäter märk-överföring med stor känslighet.
En tvåfaldig igenkänningsstrategi
Strukturerna visar att UBA6 känner igen sin nyckelpartner E2, kallad UBE2Z, med ett ”tvåhandsgrepp”. Den ena handen är en domän i slutet av UBA6 som fungerar som en dockningsyta för E2:or, medan den andra är ett katalytiskt område som sitter närmare reaktionsstället. Till skillnad från UBA1, som i huvudsak förlitar sig på dockningsregionen för att välja sina partner, använder UBA6 båda regionerna tillsammans för att omsluta UBE2Z på ett sätt som exakt alignerar de två reaktiva ytor som behövs för märk‑överföring. Extra loopsegment på UBE2Z, frånvarande i många andra E2:or, passar in i en förstorad fåra i UBA6, vilket möjliggör ett tätt men flexibelt engagemang. Om antingen dockningsytan eller det katalytiska vecket förändras misslyckas UBA6 med att ladda UBE2Z effektivt, vilket visar att båda ”händerna” krävs för korrekt partnerval.
En liten kofaktor och en flexibel strömbrytare
En överraskning är att en liten molekyl kallad inositolhexakisfosfat ligger i en ficka nära UBA6:s katalytiska veck. Denna kofaktor hjälper till att hålla fåran öppen i en konfiguration som välkomnar de ovanliga looparna hos klass IV E2:or, inklusive UBE2Z och BIRC6, samtidigt som samma region görs mindre kompatibel med de E2:or som föredras av UBA1. Förstorade strukturer av ett intermediärt tillstånd, där UBA6 och UBE2Z är bundna samtidigt till två kopior av antingen ubiquitin eller FAT10, visar hur märkningen överlämnas. En rörlig loop i UBE2Z svänger åt sidan som en grind och exponerar sin reaktiva cystein så att märkningen kan överföras. Kontaktytan kan anpassa sig: den bildar en mer polar, laddningsbaserad yta med FAT10 och en mer hydrofob, oljelik yta med ubiquitin, vilket gör det möjligt för samma E2 att hantera två kemiskt distinkta märkningar.
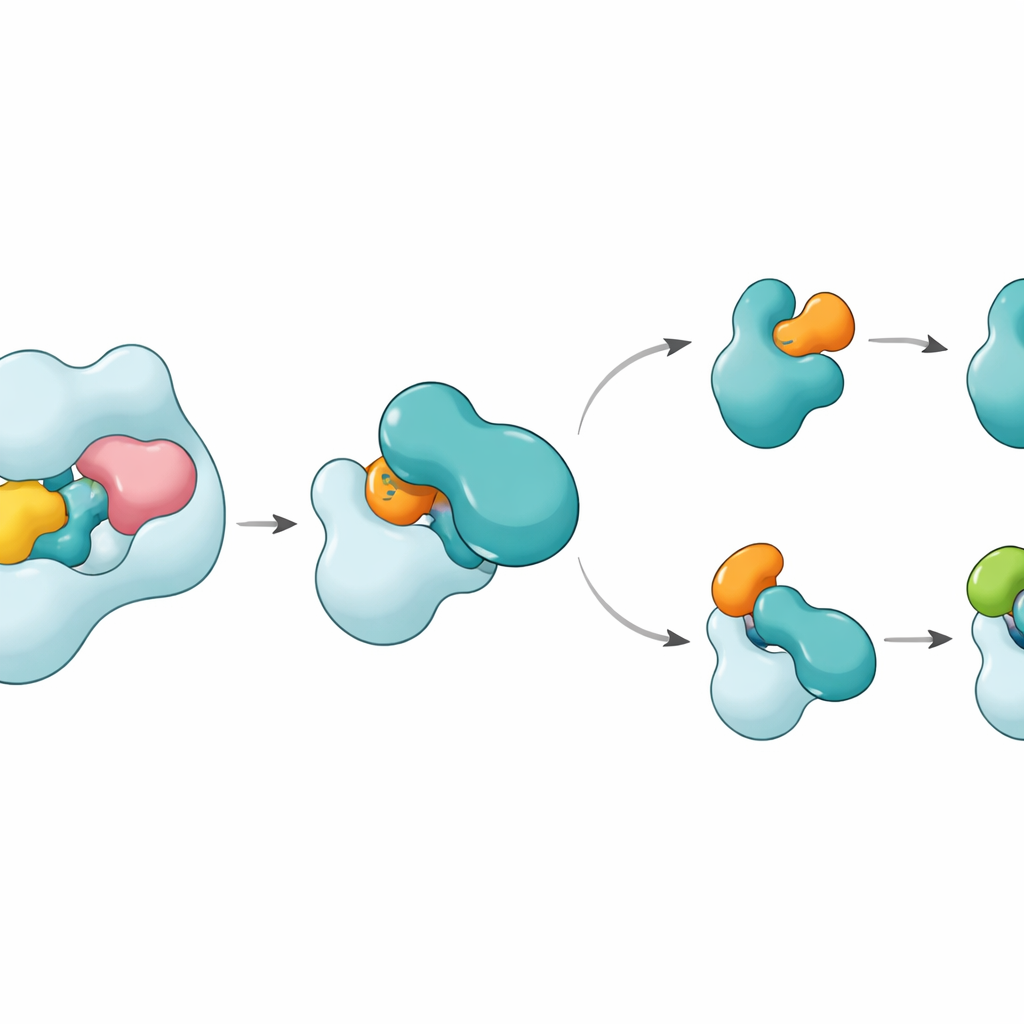
Hur detta håller cellens märkning på rätt spår
Enkelt uttryckt visar detta arbete att UBA6 har utvecklat en specialiserad, tvådelad igenkänningskula, förstärkt av en liten kofaktor, som låter det samarbeta nära med en utvald grupp E2-partners och stödja både ubiquitin- och FAT10-märkning. UBA1, däremot, använder en mer traditionell, enregionsstrategi och samarbetar med en annan uppsättning E2:or. Dessa skilda designlösningar skapar två parallella men isolerade märkningsgrenar inne i cellen: en som hanterar vardaglig proteinkontroll och en annan som kopplar proteinbortskaffning till immunsignaler. Att förstå denna arkitektoniska logik klargör inte bara hur celler upprätthåller ordning i sina proteinnätverk utan kan också vägleda framtida insatser att rikta in sig på specifika grenar av märkningssystemet vid cancer, immunitet och andra sjukdomar.
Citering: Nayak, D., Jia, L., dos Santos Bury, P. et al. Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer. Nat Commun 17, 3302 (2026). https://doi.org/10.1038/s41467-026-69882-3
Nyckelord: ubiquitin, FAT10, UBA6, proteinnedbrytning, cryo-EM