Clear Sky Science · de
Cryo‑EM-Strukturen von UBA6 zeigen Mechanismen der E1–E2‑Spezifität und des dualen Thioester‑Transfers von FAT10/Ubikuitin
Wie Zellen entscheiden, welche Proteine markiert werden
Im Inneren jeder Zelle entscheidet ein verborgenes Markierungssystem, welche Proteine recycelt, welche umgebaut und welche zur Auslösung einer Immunantwort eingesetzt werden. Zwei winzige Markierungen, Ubiquitin und FAT10, helfen bei diesen Entscheidungen und müssen mit höchster Präzision auf Trägerenzyme geladen werden. Dieser Artikel untersucht, wie ein zentrales Ladeenzym, genannt UBA6, selektiv mit bestimmten Partnerenzymen zusammenarbeitet und sowohl Ubiquitin als auch FAT10 verarbeitet. Die Ergebnisse offenbaren Gestaltungsprinzipien, die das proteinkontrollierende System der Zelle genau und anpassungsfähig halten.
Zwei parallele Markierungsstraßen
Zellen nutzen molekulare Markierungen, um Proteine für unterschiedliche Schicksale zu kennzeichnen – vom Abbau bis zu Veränderungen von Aktivität oder Lokalisation. Ubiquitin ist die bekannteste Markierung und wird in vielen zellulären Prozessen verwendet, während FAT10 spezialisierter ist und stark mit Immunantworten und Entzündungen verknüpft wird. Beide Marker werden über eine dreistufige Übergabe durch E1-, E2- und E3‑Enzyme angeheftet. Die E1‑Enzyme übernehmen den ersten, energieverbrauchenden Schritt, aktivieren die Markierung und geben sie an E2s weiter, die dann mit E3s zusammenarbeiten, um die Markierung an Zielproteine zu hängen. Menschen sind ungewöhnlich darin, zwei E1‑Enzyme für Ubiquitin zu besitzen: UBA1, das den klassischen Ubiquitinweg bedient, und UBA6, das nicht nur Ubiquitin, sondern auch FAT10 aktiviert und damit eine Brücke zwischen alltäglicher Proteinkontrolle und immunreguliertem Proteinabbau bildet.
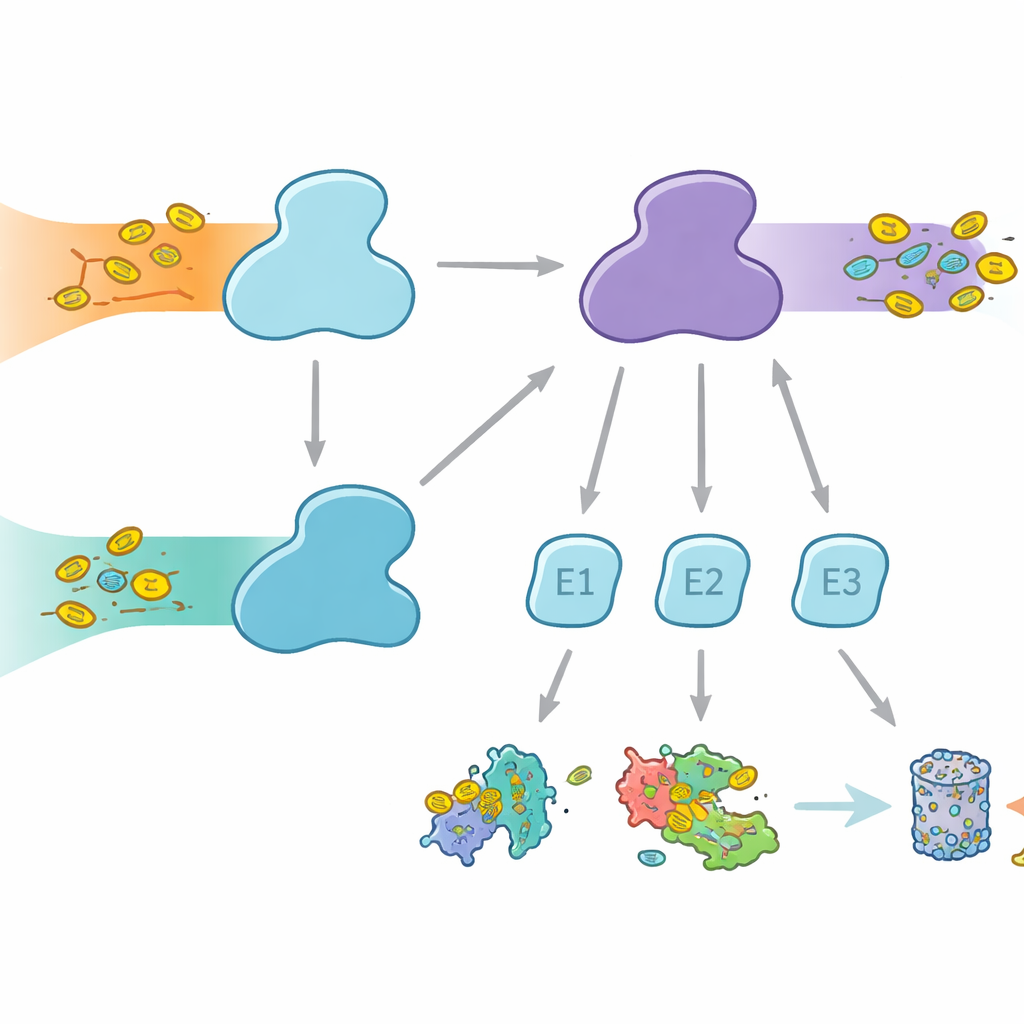
Warum die Wahl des Partners wichtig ist
Obwohl UBA1 und UBA6 oberflächlich ähnlich aussehen, arbeiten sie mit weitgehend unterschiedlichen Sätzen von E2‑Partnerenzymen und schaffen so parallele Verzweigungen des Markierungssystems. Manche E2s können mit beiden E1s zusammenarbeiten, andere sind exklusive Partner. Diese Selektivität ist entscheidend: Würden falsche Kombinationen entstehen, könnten immunverknüpfte FAT10‑Signale in gewöhnliche Ubiquitinwege „auslaufen“ oder umgekehrt, was zelluläre Entscheidungen durcheinanderbringen würde. Bislang war unklar, wie UBA6 seine bevorzugten E2s erkennen kann, insbesondere eine spezialisierte Gruppe, die als Klasse‑IV‑E2s bekannt ist, während UBA1 sie weitgehend ignoriert. Die Autoren machten sich daran, UBA6 und seine Partner im Moment der Aktion mit hochauflösender Kryo‑Elektronenmikroskopie und biochemischen Experimenten, die die Markierungsübertragung sehr empfindlich messen, einzufangen.
Eine Erkennungsstrategie mit zwei Griffen
Die Strukturen zeigen, dass UBA6 seinen Schlüsselpartner E2, genannt UBE2Z, mit einem „zwei‑händigen“ Griff erkennt. Eine Hand ist eine Domäne am Ende von UBA6, die als Andockstelle für E2s dient, die andere ist eine katalytische Region, die näher an der Reaktionsstelle sitzt. Anders als UBA1, das hauptsächlich auf die Andockregion zur Partnerwahl setzt, nutzt UBA6 beide Regionen zusammen, um UBE2Z so zu umschließen, dass die beiden reaktiven Stellen für die Markierungsübertragung präzise ausgerichtet sind. Zusätzliche Schleifenabschnitte auf UBE2Z, die bei vielen anderen E2s fehlen, passen in eine vergrößerte Rinne in UBA6 und erlauben ein enges, zugleich flexibles Andocken. Werden entweder die Andockfläche oder die katalytische Kluft verändert, kann UBA6 UBE2Z nicht effizient beladen, was zeigt, dass beide „Hände" für die korrekte Partnerwahl erforderlich sind.
Ein kleiner Cofaktor und ein flexibler Schalter
Überraschend ist, dass ein winziges Molekül, Inositolhexakisphosphat, in eine Tasche nahe der katalytischen Kluft von UBA6 eingeschmiegt ist. Dieser Cofaktor hilft, die Rinne in einer Konfiguration offen zu halten, die die ungewöhnlichen Schleifen der Klasse‑IV‑E2s, einschließlich UBE2Z und BIRC6, willkommen heißt, während er dieselbe Region weniger kompatibel für von UBA1 bevorzugte E2s macht. Nahaufnahmen eines Zwischenzustands, in dem UBA6 und UBE2Z gleichzeitig an zwei Kopien entweder von Ubiquitin oder FAT10 gebunden sind, zeigen, wie die Markierung übergeben wird. Eine bewegliche Schleife in UBE2Z schwenkt wie ein Tor zur Seite und legt ihr reaktives Cystein frei, sodass die Markierung übertragen werden kann. Die Kontaktfläche ist anpassungsfähig: Sie bildet mit FAT10 eine polarere, ladungsbasierte Schnittstelle und mit Ubiquitin eine hydrophobere, „ölartige“ Oberfläche, wodurch derselbe E2 zwei chemisch unterschiedliche Marker handhaben kann.
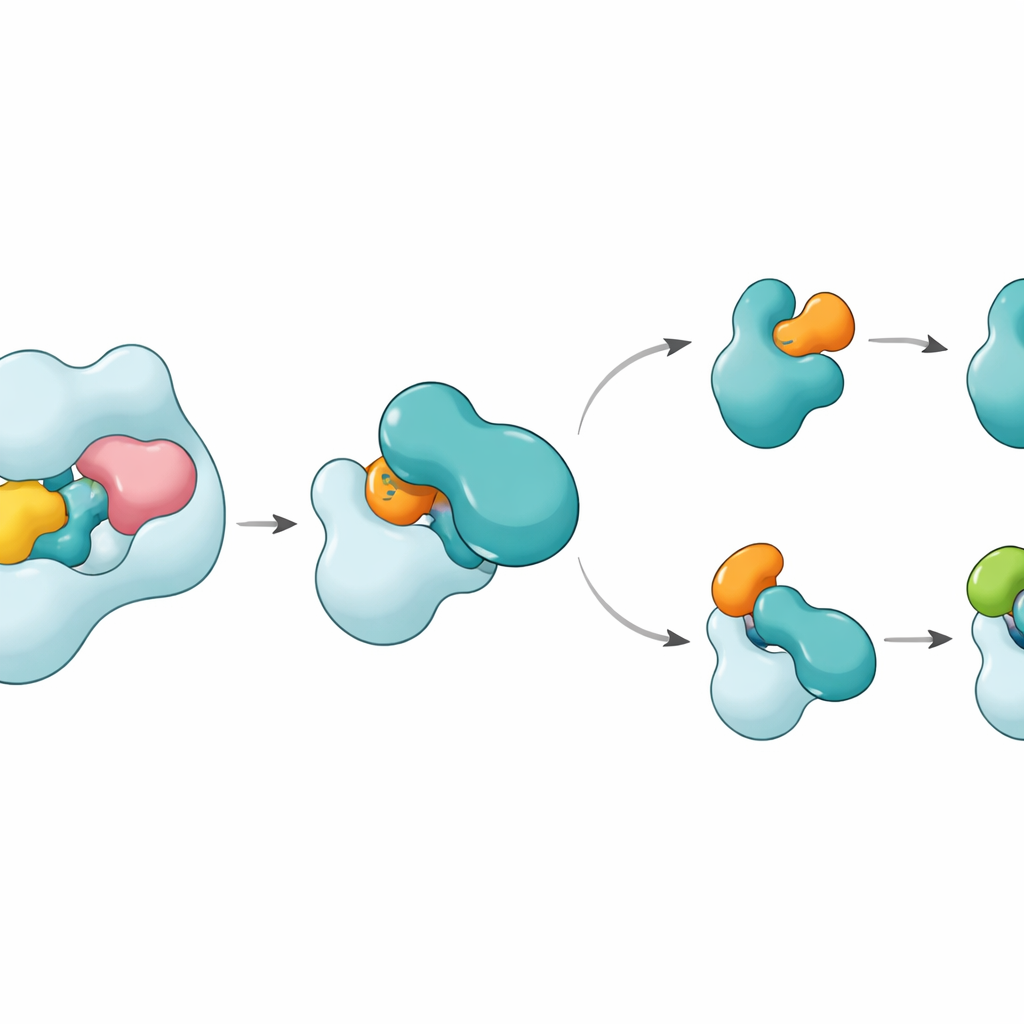
Wie das die zelluläre Markierung auf Kurs hält
Vereinfacht zeigt diese Arbeit, dass UBA6 eine spezialisierte, zweiteilige Erkennungswiege entwickelt hat, die durch einen kleinen Cofaktor verstärkt wird und es ihm ermöglicht, eng mit einer ausgewählten Gruppe von E2‑Partnern zu arbeiten und sowohl Ubiquitin‑ als auch FAT10‑Markierungen zu unterstützen. UBA1 hingegen verwendet eine eher traditionelle, einregionale Strategie und arbeitet mit einem anderen E2‑Satz zusammen. Diese unterschiedlichen Gestaltungsansätze schaffen zwei parallele, aber voneinander isolierte Markierungszweige in der Zelle: einen für die alltägliche Proteinkontrolle und einen, der die Proteinentsorgung an Immun‑Signale koppelt. Das Verständnis dieser architektonischen Logik klärt nicht nur, wie Zellen Ordnung in ihren Proteinnetzwerken bewahren, sondern kann auch künftige Bemühungen informieren, spezifische Zweige des Markierungssystems in Krebs, Immunität und anderen Erkrankungen gezielt anzugehen.
Zitation: Nayak, D., Jia, L., dos Santos Bury, P. et al. Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer. Nat Commun 17, 3302 (2026). https://doi.org/10.1038/s41467-026-69882-3
Schlüsselwörter: Ubiquitin, FAT10, UBA6, Proteinabbau, Cryo‑EM