Clear Sky Science · es
Estructuras por crio‑EM de UBA6 revelan los mecanismos de especificidad E1–E2 y la transferencia tioéster dual de FAT10/ubiquitina
Cómo deciden las células qué proteínas etiquetar
Dentro de cada célula, un sistema de etiquetado oculto decide qué proteínas se envían al reciclaje, cuáles se remodelan y cuáles ayudan a montar una respuesta inmune. Dos etiquetas diminutas, la ubiquitina y FAT10, participan en estas decisiones y deben cargarse en enzimas portadoras con precisión exquisita. Este artículo explora cómo una enzima cargadora clave, llamada UBA6, trabaja selectivamente con ciertos enzimas socias y maneja tanto ubiquitina como FAT10, revelando principios de diseño que mantienen el sistema de control proteico celular preciso y adaptable.
Dos autopistas de etiquetado en paralelo
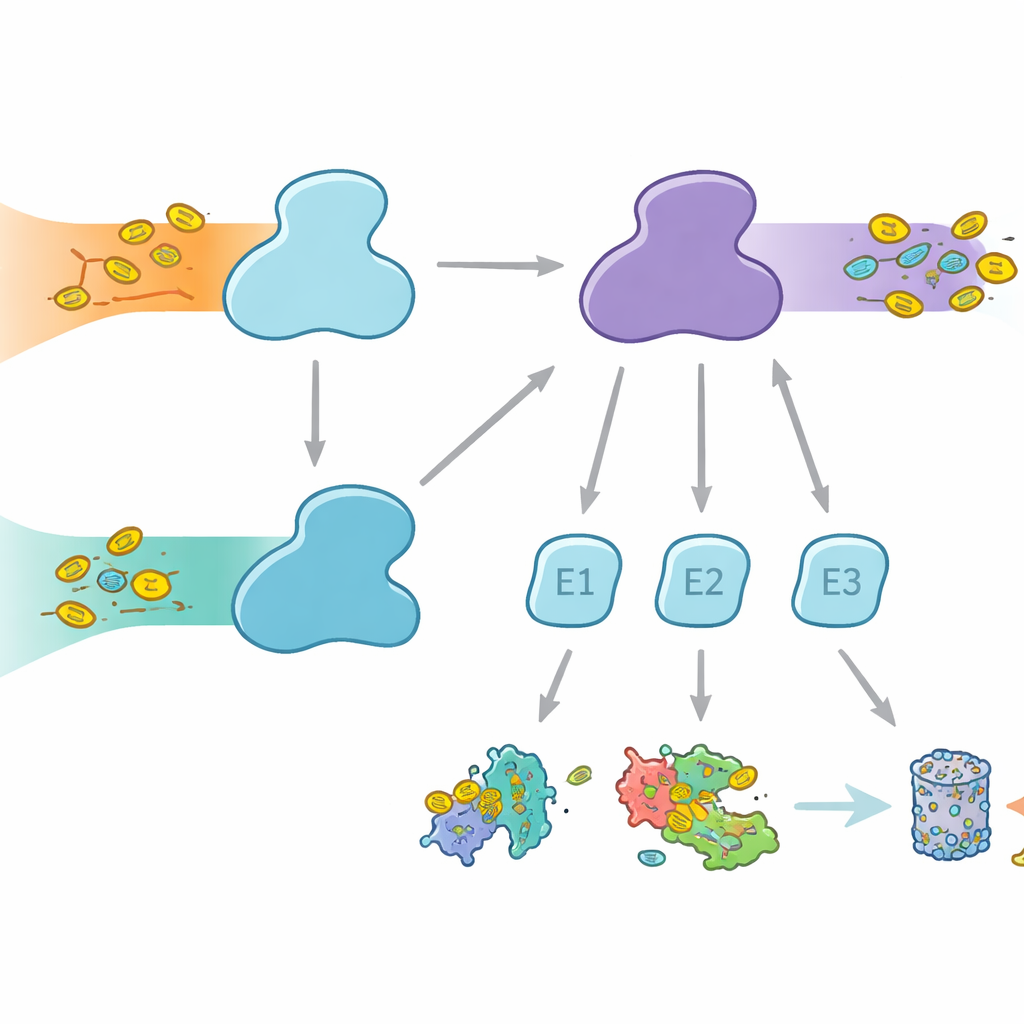
Las células usan etiquetas moleculares para marcar proteínas con distintos destinos, desde la degradación hasta cambios en la actividad o la localización. La ubiquitina es la etiqueta más conocida, empleada en numerosos procesos celulares, mientras que FAT10 es más especializada y está fuertemente ligada a respuestas inmunes e inflamación. Ambas etiquetas se unen mediante un relevo de tres pasos que involucra enzimas E1, E2 y E3. Las E1 realizan el primer paso, que consume energía, activando la etiqueta y pasándola a las E2, que a su vez colaboran con las E3 para adjuntar la etiqueta a las proteínas diana. Los humanos son inusuales por tener dos enzimas E1 para la ubiquitina: UBA1, que sirve la vía clásica de la ubiquitina, y UBA6, que no solo activa ubiquitina sino también FAT10, formando un puente entre el control proteico cotidiano y la eliminación de proteínas regulada por la inmunidad.
Por qué importa la elección de la pareja
Aunque UBA1 y UBA6 parecen, en términos generales, similares, trabajan con conjuntos en gran parte distintos de enzimas E2 socias, creando ramas paralelas del sistema de etiquetado. Algunas E2 pueden funcionar con ambas E1, pero otras son parejas exclusivas. Esta selectividad es vital: si se formaran combinaciones incorrectas, las señales de FAT10 relacionadas con la inmunidad podrían filtrarse en las vías ordinarias de ubiquitina, o viceversa, desbaratando las decisiones celulares. Hasta ahora no quedaba claro cómo UBA6 podía reconocer a sus E2 preferidas, en especial a un grupo especializado conocido como E2 de clase IV, mientras que UBA1 los ignora en gran medida. Los autores se propusieron capturar a UBA6 y sus socios en acción mediante crio‑microscopía electrónica de alta resolución y experimentos bioquímicos que miden la transferencia de etiquetas con gran sensibilidad.
Una estrategia de reconocimiento de doble agarre
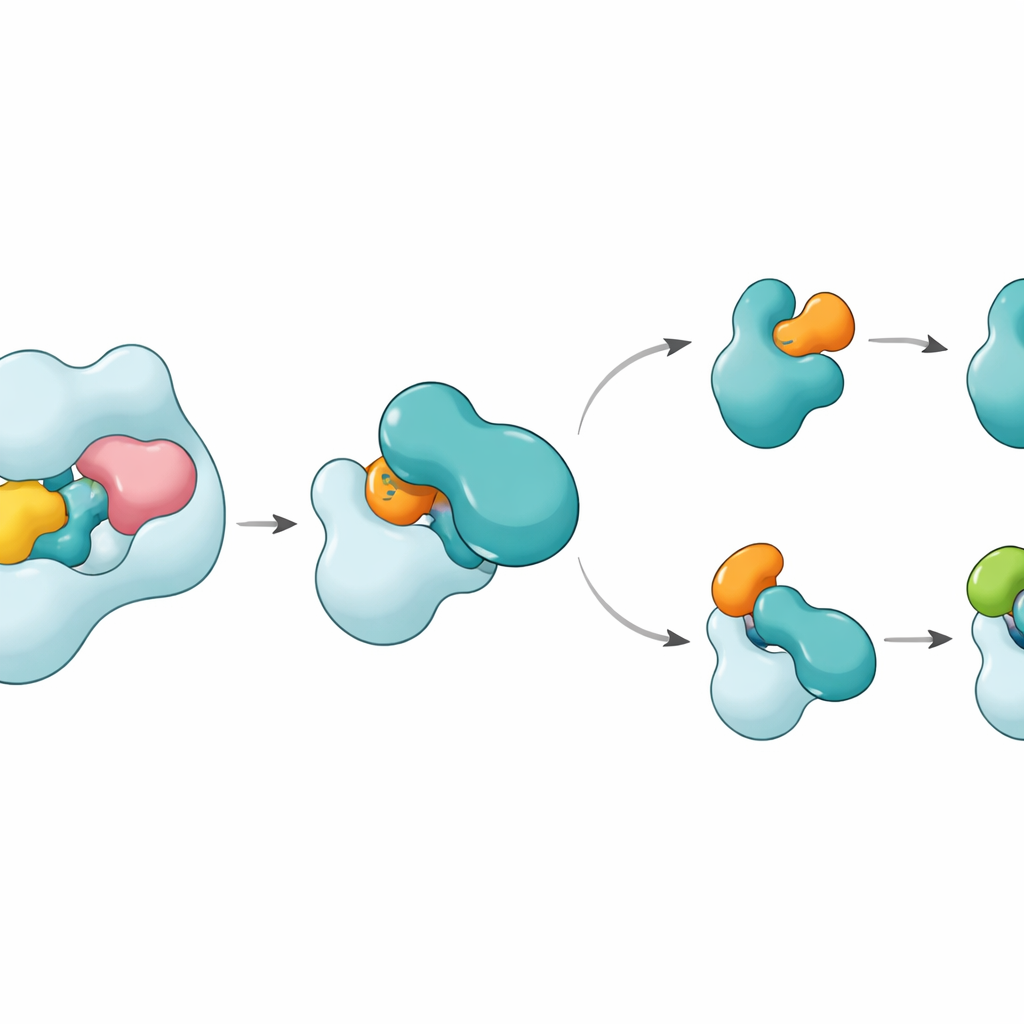
Las estructuras revelan que UBA6 reconoce a su socio E2 clave, denominado UBE2Z, usando un “agarre de dos manos”. Una mano es un dominio en el extremo de UBA6 que actúa como muelle de acoplamiento para las E2, mientras que la otra es una región catalítica que se sitúa más cerca del sitio de la reacción química. A diferencia de UBA1, que se apoya principalmente en la región de acoplamiento para seleccionar a sus socios, UBA6 utiliza ambas regiones juntas para abrazar a UBE2Z de forma que alinee con precisión los dos sitios reactivos necesarios para la transferencia de la etiqueta. Segmentos de bucle adicionales en UBE2Z, ausentes en muchas otras E2, encajan en una ranura agrandada en UBA6, permitiendo un compás estrecho pero flexible. Si se altera cualquiera de la superficie de acoplamiento o la hendidura catalítica, UBA6 no consigue cargar eficientemente a UBE2Z, lo que muestra que ambas “manos” son necesarias para la elección correcta de la pareja.
Un cofactor pequeño y un interruptor flexible
Una sorpresa es que una molécula diminuta llamada inositol hexakisfosfato se aloja en un bolsillo cercano a la hendidura catalítica de UBA6. Este cofactor ayuda a mantener la ranura abierta en una configuración que acoge los bucles inusuales de las E2 de clase IV, incluyendo UBE2Z y BIRC6, mientras hace que la misma región sea menos compatible con las E2 favorecidas por UBA1. Estructuras ampliadas de un estado intermedio, en el que UBA6 y UBE2Z están unidos simultáneamente a dos copias ya sea de ubiquitina o de FAT10, muestran cómo se realiza la entrega de la etiqueta. Un bucle móvil en UBE2Z se desplaza como una puerta, exponiendo su cisteína reactiva para que se transfiera la etiqueta. La superficie de contacto puede adaptarse: forma una interfaz más polar, basada en cargas, con FAT10 y una interfaz más hidrofóbica, tipo aceite, con ubiquitina, permitiendo que la misma E2 maneje dos etiquetas químicamente distintas.
Cómo esto mantiene el etiquetado celular en camino
En términos sencillos, este trabajo muestra que UBA6 ha evolucionado una cuna de reconocimiento especializada de dos partes, reforzada por un pequeño cofactor, que le permite trabajar estrechamente con un grupo selecto de socios E2 y soportar tanto el etiquetado con ubiquitina como con FAT10. UBA1, en contraste, utiliza una estrategia más tradicional basada en una sola región y se asocia con un conjunto distinto de E2. Estas soluciones de diseño distintas crean dos ramas paralelas pero aisladas de etiquetado dentro de la célula: una que gestiona el control proteico cotidiano y otra que enlaza la eliminación de proteínas con señales inmunitarias. Comprender esta lógica arquitectónica no solo aclara cómo las células mantienen el orden en sus redes proteicas, sino que también puede orientar futuros esfuerzos para dirigirse a ramas específicas del sistema de etiquetado en cáncer, inmunidad y otras enfermedades.
Cita: Nayak, D., Jia, L., dos Santos Bury, P. et al. Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer. Nat Commun 17, 3302 (2026). https://doi.org/10.1038/s41467-026-69882-3
Palabras clave: ubiquitina, FAT10, UBA6, degradación de proteínas, crio‑EM