Clear Sky Science · en
Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer
How Cells Decide Which Proteins to Tag
Inside every cell, a hidden tagging system decides which proteins are sent for recycling, which are remodeled, and which help mount an immune response. Two tiny tags, ubiquitin and FAT10, help make these decisions, and they must be loaded onto carrier enzymes with exquisite precision. This paper explores how one key loader enzyme, called UBA6, selectively works with certain partner enzymes and handles both ubiquitin and FAT10, revealing design principles that keep the cell’s protein-control system accurate and adaptable.
Two Parallel Tagging Highways
Cells use molecular tags to mark proteins for different fates, from degradation to changes in activity or location. Ubiquitin is the best-known tag, used across many cellular processes, while FAT10 is more specialized and strongly linked to immune responses and inflammation. Both tags are attached through a three-step relay involving E1, E2, and E3 enzymes. The E1 enzymes perform the first, energy‑consuming step, activating the tag and passing it to E2s, which then work with E3s to attach the tag to target proteins. Humans are unusual in having two E1 enzymes for ubiquitin: UBA1, which serves the classic ubiquitin pathway, and UBA6, which not only activates ubiquitin but also FAT10, forming a bridge between everyday protein control and immune‑regulated protein disposal.
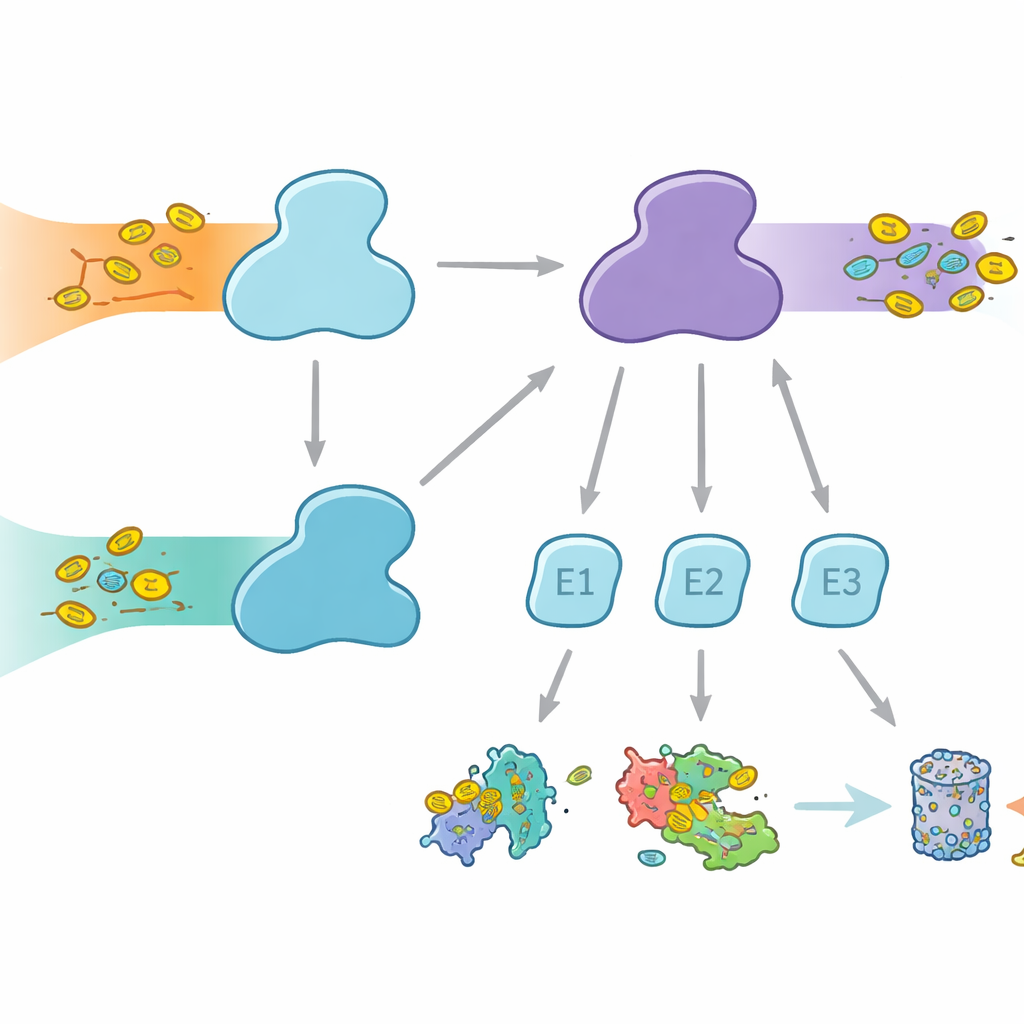
Why Partner Choice Matters
Even though UBA1 and UBA6 look broadly similar, they work with largely different sets of E2 partner enzymes, creating parallel branches of the tagging system. Some E2s can work with both E1s, but others are exclusive partners. This selectivity is vital: if the wrong combinations formed, immune‑linked FAT10 signals could leak into ordinary ubiquitin pathways, or vice versa, scrambling cellular decisions. Until now, it was unclear how UBA6 could recognize its own preferred E2s, especially a specialized group known as class IV E2s, while UBA1 largely ignores them. The authors set out to capture UBA6 and its partners in the act using high‑resolution cryo‑electron microscopy and biochemical experiments that measure tag transfer with great sensitivity.
A Dual-Grip Recognition Strategy
The structures reveal that UBA6 recognizes its key partner E2, called UBE2Z, using a “two‑handed” grip. One hand is a domain at the end of UBA6 that serves as a docking dock for E2s, while the other is a catalytic region that sits closer to the chemical reaction site. Unlike UBA1, which relies mostly on the docking region to select its partners, UBA6 uses both regions together to cradle UBE2Z in a way that precisely aligns the two reactive sites needed for tag transfer. Extra loop segments on UBE2Z, absent in many other E2s, fit into an enlarged groove in UBA6, allowing tight yet flexible engagement. If either the docking surface or the catalytic cleft is altered, UBA6 fails to load UBE2Z efficiently, showing that both “hands” are required for correct partner choice.
A Small Cofactor and a Flexible Switch
One surprise is that a tiny molecule called inositol hexakisphosphate nestles into a pocket near UBA6’s catalytic cleft. This cofactor helps hold the groove open in a configuration that welcomes the unusual loops of class IV E2s, including UBE2Z and BIRC6, while making the same region less compatible with E2s favored by UBA1. Zoomed‑in structures of an intermediate state, where UBA6 and UBE2Z are simultaneously bound to two copies of either ubiquitin or FAT10, show how the tag is handed off. A movable loop in UBE2Z swings aside like a gate, exposing its reactive cysteine so the tag can be transferred. The contact surface can adapt: it forms a more polar, charge‑based interface with FAT10 and a more hydrophobic, oil‑like interface with ubiquitin, allowing the same E2 to handle two chemically distinct tags.
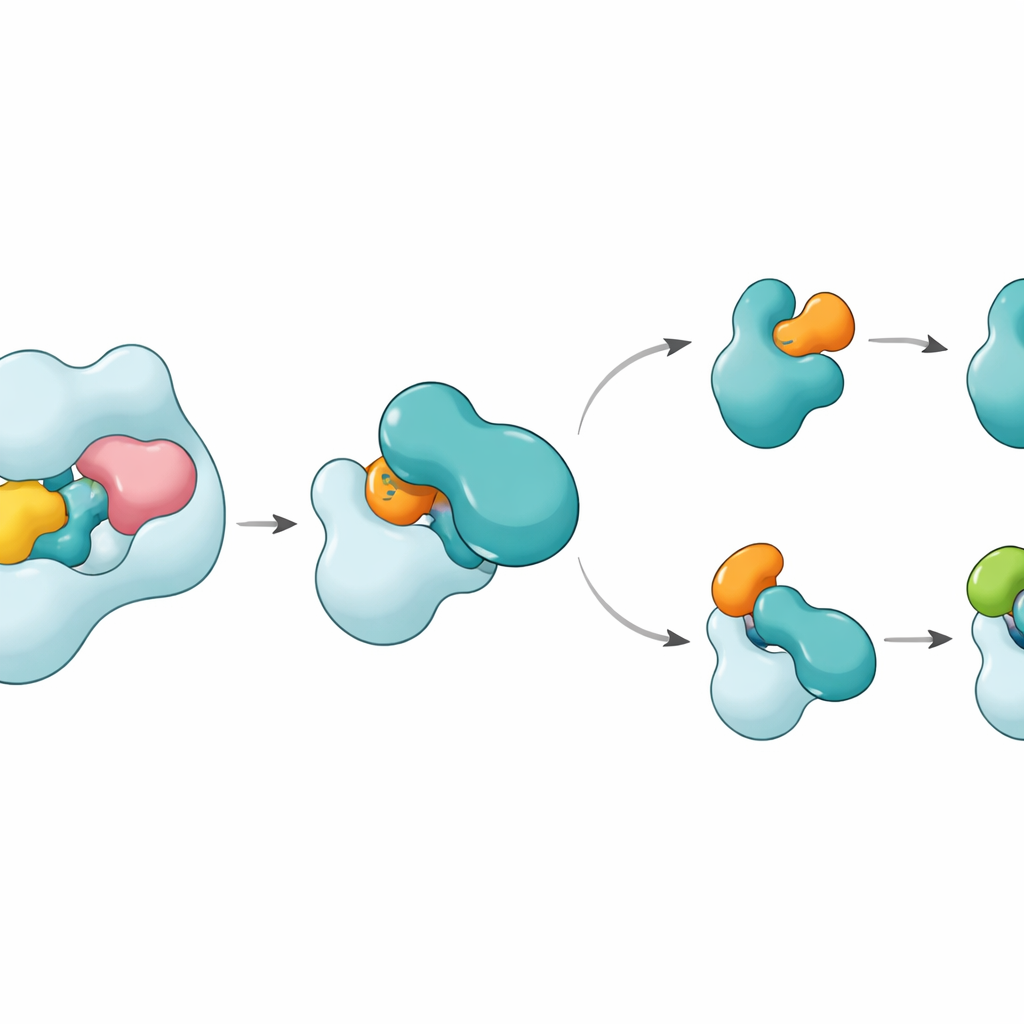
How This Keeps Cellular Tagging on Track
In simple terms, this work shows that UBA6 has evolved a specialized, two‑part recognition cradle, reinforced by a small cofactor, that lets it work closely with a select group of E2 partners and support both ubiquitin and FAT10 tagging. UBA1, in contrast, uses a more traditional, single‑region strategy and partners with a different E2 set. These distinct design solutions create two parallel but insulated tagging branches inside the cell: one handling everyday protein control, the other linking protein disposal to immune signals. Understanding this architectural logic not only clarifies how cells maintain order in their protein networks but may also inform future efforts to target specific branches of the tagging system in cancer, immunity, and other diseases.
Citation: Nayak, D., Jia, L., dos Santos Bury, P. et al. Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer. Nat Commun 17, 3302 (2026). https://doi.org/10.1038/s41467-026-69882-3
Keywords: ubiquitin, FAT10, UBA6, protein degradation, cryo-EM