Clear Sky Science · pl
Struktury cryo-EM UBA6 ujawniają mechanizmy swoistości E1–E2 oraz podwójnego transferu tioestrycznego FAT10/ubiquityny
Jak komórki decydują, które białka oznaczać
W każdej komórce działa ukryty system tagowania, który decyduje, jakie białka trafiają do recyklingu, które są przebudowywane, a które wspierają odpowiedź immunologiczną. Dwa niewielkie znaczniki, ubiquityna i FAT10, pomagają podejmować te decyzje i muszą być precyzyjnie załadowane na enzymy nośnikowe. W artykule tym opisano, jak jeden kluczowy enzym ładujący, zwany UBA6, selektywnie współpracuje z określonymi enzymami partnerami i obsługuje zarówno ubiquitynę, jak i FAT10, ujawniając zasady konstrukcyjne, które utrzymują system kontroli białek w komórce dokładnym i elastycznym.
Dwie równoległe autostrady tagowania
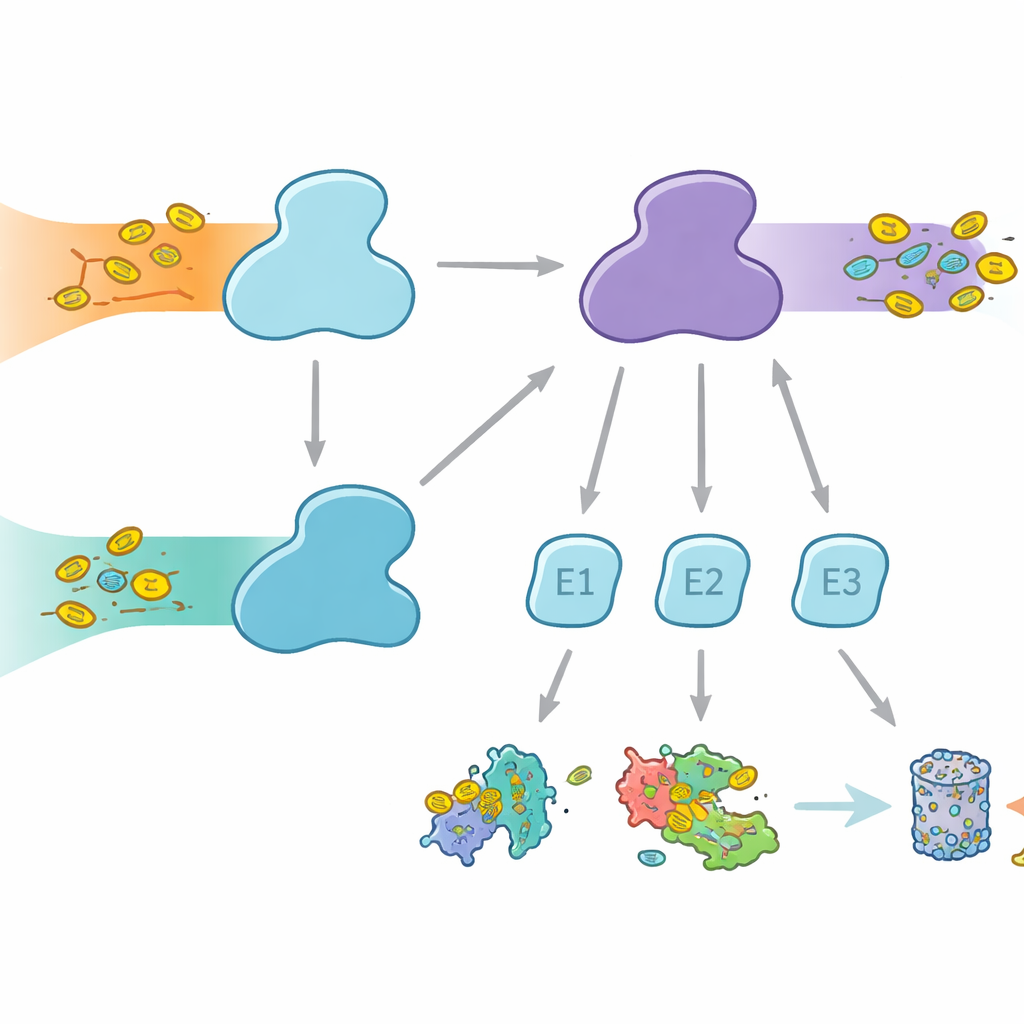
Komórki używają molekularnych znaczników do oznaczania białek w różnych celach — od degradacji po zmiany aktywności lub lokalizacji. Ubiquityna jest najlepiej znanym znacznikiem, używanym w wielu procesach komórkowych, natomiast FAT10 ma bardziej wyspecjalizowaną rolę i jest silnie powiązana z reakcjami immunologicznymi i stanami zapalnymi. Oba znaczniki są przyłączane w trójetapowym relayu obejmującym enzymy E1, E2 i E3. Enzymy E1 wykonują pierwszy, energetyczny etap, aktywując znacznik i przekazując go do E2, które następnie współpracują z E3, aby przyłączyć znacznik do białek docelowych. Ludzie są nietypowi pod tym względem, że mają dwa enzymy E1 dla ubiquityny: UBA1, obsługujący klasyczną ścieżkę ubiquitynacji, oraz UBA6, który nie tylko aktywuje ubiquitynę, lecz także FAT10, tworząc pomost między codzienną kontrolą białek a immunologicznie regulowaną eliminacją białek.
Dlaczego wybór partnera ma znaczenie
Chociaż UBA1 i UBA6 są ogólnie podobne, współpracują głównie z różnymi zestawami enzymów E2, tworząc równoległe gałęzie systemu tagowania. Niektóre E2 mogą współdziałać z oboma E1, ale inne są ekskluzywnymi partnerami. Ta selektywność jest kluczowa: gdyby powstawały niewłaściwe kombinacje, sygnały FAT10 związane z układem immunologicznym mogłyby przenikać do zwykłych szlaków ubiquitynacji lub odwrotnie, zaburzając decyzje komórkowe. Dotąd nie było jasne, jak UBA6 rozpoznaje swoje preferowane E2, zwłaszcza wyspecjalizowaną grupę znaną jako klasy IV E2, podczas gdy UBA1 w dużej mierze je pomija. Autorzy postanowili uchwycić UBA6 i jego partnerów w akcji, wykorzystując wysokorozdzielczą krio‑elektronową mikroskopię oraz eksperymenty biochemiczne mierzące transfer znacznika z dużą czułością.
Strategia rozpoznawania w podwójnym ujęciu
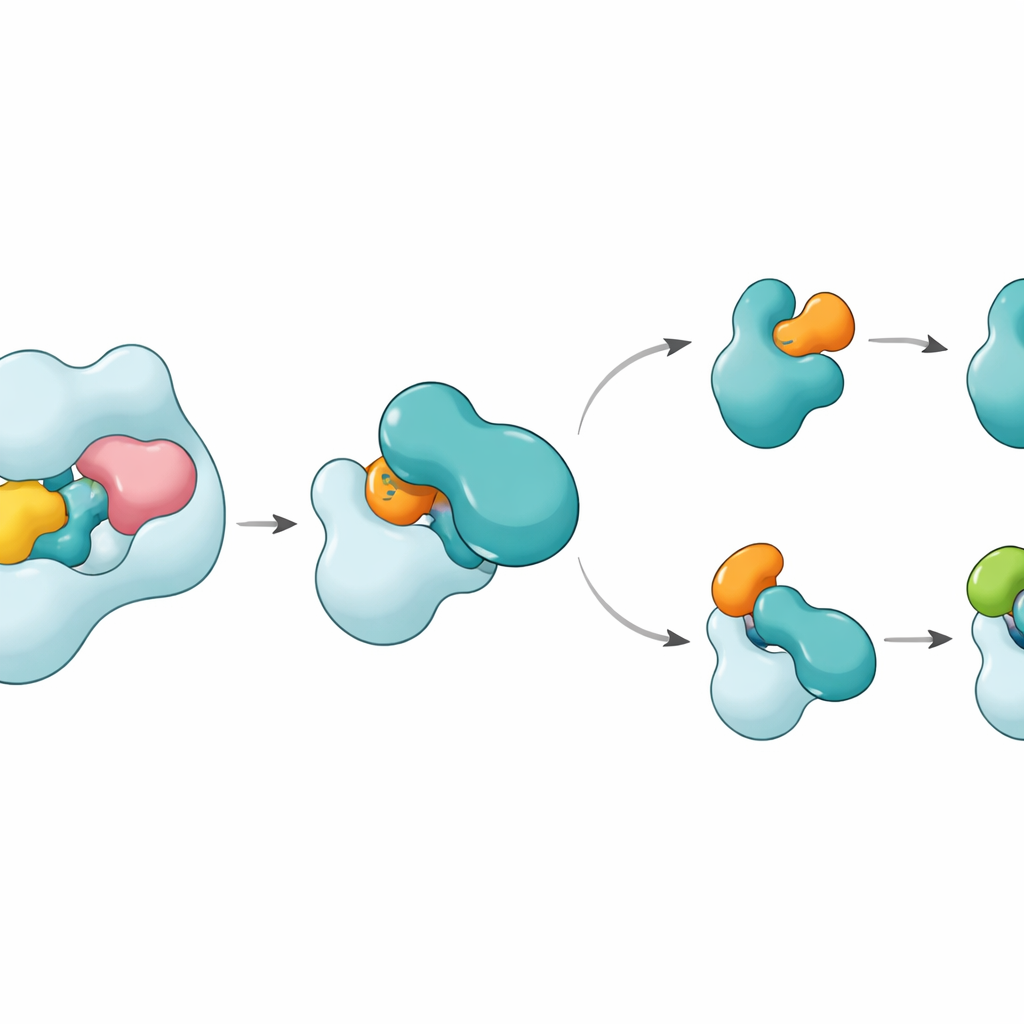
Struktury pokazują, że UBA6 rozpoznaje kluczowego partnera E2, zwanego UBE2Z, przy użyciu „dwuręcznego” chwytu. Jedna „ręka” to domena na końcu UBA6, służąca jako dok dla E2, a druga to region katalityczny położony bliżej miejsca reakcji chemicznej. W odróżnieniu od UBA1, które opiera się głównie na regionie dokującym przy wyborze partnerów, UBA6 używa obu regionów jednocześnie, obejmując UBE2Z w sposób, który precyzyjnie wyrównuje dwa reaktywne miejsca potrzebne do transferu znacznika. Dodatkowe pętle na UBE2Z, nieobecne w wielu innych E2, mieszczą się w powiększonej szczelinie UBA6, umożliwiając ścisłe, ale elastyczne wiązanie. Jeśli zmienione zostanie któreś z miejsc — powierzchnia dokująca lub szczelina katalityczna — UBA6 nie ładuje efektywnie UBE2Z, co pokazuje, że obie „ręce” są potrzebne do właściwego wyboru partnera.
Mały kofaktor i giętki przełącznik
Jednym z zaskoczeń jest stwierdzenie, że niewielka cząsteczka zwana inozytolheksakisfosforanem zagnieżdża się w kieszeni w pobliżu szczeliny katalitycznej UBA6. Ten kofaktor pomaga utrzymać szczelinę otwartą w konfiguracji sprzyjającej przyjęciu nietypowych pętli klasy IV E2, w tym UBE2Z i BIRC6, jednocześnie czyniąc ten sam region mniej kompatybilnym z E2 preferowanymi przez UBA1. Strukturami zbliżonymi do stanu pośredniego, gdzie UBA6 i UBE2Z są jednocześnie związane z dwoma kopiami albo ubiquityny, albo FAT10, pokazano, jak następuje przekazanie znacznika. Ruchoma pętla w UBE2Z odchyla się niczym bramka, odsłaniając reaktywną cysteinę, aby możliwy był transfer znacznika. Powierzchnia kontaktu potrafi się dostosować: z FAT10 tworzy bardziej polarny, oparty na ładunkach interfejs, a z ubiquityną bardziej hydrofobowy, „oleisty” interfejs, co pozwala temu samemu E2 obsługiwać dwa chemicznie różne znaczniki.
Jak to utrzymuje porządek w tagowaniu komórkowym
Mówiąc prosto, praca ta pokazuje, że UBA6 wyewoluowało wyspecjalizowaną, dwu‑częściową rozpoznawczą kołyskę, wzmocnioną przez mały kofaktor, która pozwala mu ściśle współdziałać z wybranym zestawem partnerów E2 i wspierać zarówno tagowanie ubiquityną, jak i FAT10. UBA1, przeciwnie, stosuje bardziej tradycyjną strategię opartą na pojedynczym regionie i współpracuje z innym zestawem E2. Te odmienne rozwiązania konstrukcyjne tworzą dwie równoległe, lecz izolowane gałęzie tagowania w komórce: jedna obsługująca codzienną kontrolę białek, druga łącząca usuwanie białek z sygnałami immunologicznymi. Zrozumienie tej architektonicznej logiki nie tylko wyjaśnia, jak komórki utrzymują porządek w sieciach białkowych, lecz może także ukierunkować przyszłe wysiłki mające na celu celowanie w konkretne gałęzie systemu tagowania w kontekście nowotworów, odporności i innych chorób.
Cytowanie: Nayak, D., Jia, L., dos Santos Bury, P. et al. Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer. Nat Commun 17, 3302 (2026). https://doi.org/10.1038/s41467-026-69882-3
Słowa kluczowe: ubiquityna, FAT10, UBA6, degradacja białek, cryo-EM