Clear Sky Science · it
Strutture Cryo-EM di UBA6 rivelano i meccanismi della specificità E1–E2 e il trasferimento tiolestere duale FAT10/ubiquitina
Come le cellule decidono quali proteine taggare
All’interno di ogni cellula, un sistema di etichettatura nascosto decide quali proteine vengono inviate al riciclo, quali vengono rimodellate e quali contribuiscono alla risposta immunitaria. Due piccole etichette, ubiquitina e FAT10, guidano queste decisioni e devono essere caricate su enzimi trasportatori con precisione estrema. Questo articolo esplora come un enzima caricatore chiave, chiamato UBA6, lavori selettivamente con certi enzimi partner e gestisca sia ubiquitina sia FAT10, rivelando principi di progetto che mantengono il sistema di controllo proteico cellulare accurato e adattabile.
Due autostrade parallele di etichettatura
Le cellule usano etichette molecolari per segnare le proteine destinandole a destini diversi, dalla degradazione a cambiamenti di attività o localizzazione. L’ubiquitina è l’etichetta più nota, impiegata in molti processi cellulari, mentre FAT10 è più specializzata ed è fortemente collegata alle risposte immunitarie e all’infiammazione. Entrambe le etichette vengono attaccate attraverso un trasferimento in tre passaggi che coinvolge gli enzimi E1, E2 ed E3. Gli E1 svolgono il primo passaggio che richiede energia, attivando l’etichetta e trasferendola agli E2, i quali poi collaborano con gli E3 per attaccare l’etichetta alle proteine bersaglio. Negli esseri umani è insolito avere due enzimi E1 per l’ubiquitina: UBA1, che serve la via classica dell’ubiquitina, e UBA6, che non solo attiva l’ubiquitina ma anche FAT10, formando un ponte tra il controllo proteico quotidiano e lo smaltimento delle proteine regolato dal sistema immunitario.
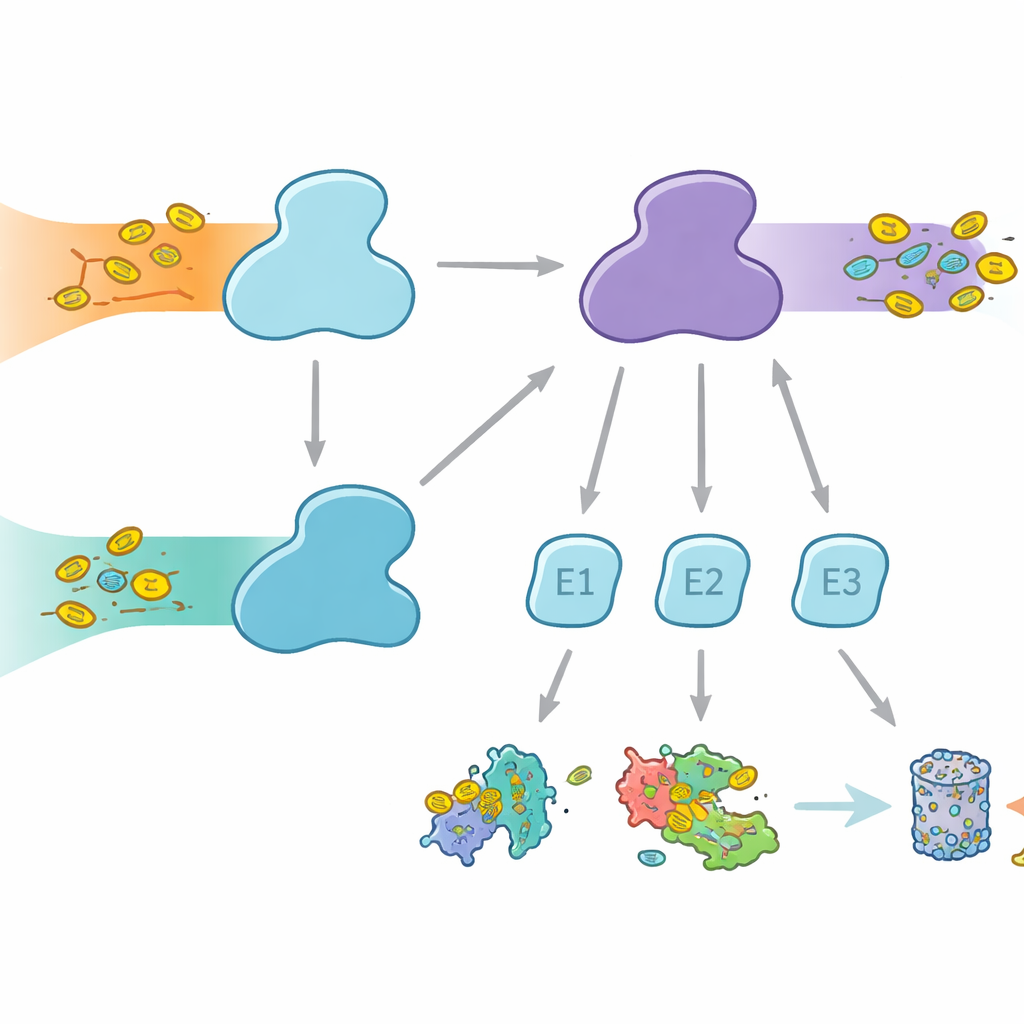
Perché la scelta del partner è importante
Pur avendo UBA1 e UBA6 un aspetto generale simile, lavorano con insiemi di enzimi E2 in gran parte differenti, creando rami paralleli del sistema di etichettatura. Alcuni E2 possono collaborare con entrambi gli E1, ma altri sono partner esclusivi. Questa selettività è vitale: se si formassero combinazioni errate, i segnali legati a FAT10 connessi all’immunità potrebbero infiltrarsi nelle vie ordinarie dell’ubiquitina, o viceversa, confondendo le decisioni cellulari. Fino ad ora non era chiaro come UBA6 potesse riconoscere i suoi E2 preferiti, in particolare un gruppo specializzato noto come E2 di classe IV, mentre UBA1 li ignora in gran parte. Gli autori hanno quindi cercato di catturare UBA6 e i suoi partner in azione usando la crio‑microscopia elettronica ad alta risoluzione e esperimenti biochimici che misurano il trasferimento delle etichette con grande sensibilità.
Strategia di riconoscimento a doppia presa
Le strutture rivelano che UBA6 riconosce il suo partner E2 principale, chiamato UBE2Z, usando una «presa a due mani». Una mano è un dominio all’estremità di UBA6 che funge da punto di ancoraggio per gli E2, mentre l’altra è una regione catalitica posta più vicino al sito della reazione chimica. Diversamente da UBA1, che si affida principalmente alla regione di ancoraggio per selezionare i partner, UBA6 usa entrambe le regioni insieme per accogliere UBE2Z in modo da allineare con precisione i due siti reattivi necessari per il trasferimento dell’etichetta. Segmenti di loop extra su UBE2Z, assenti in molti altri E2, si inseriscono in una scanalatura ampliata di UBA6, permettendo un’accoppiamento stretto ma flessibile. Se la superficie di ancoraggio o la fessura catalitica vengono alterate, UBA6 non riesce a caricare efficacemente UBE2Z, dimostrando che entrambe le «mani» sono necessarie per la corretta scelta del partner.
Un piccolo cofattore e un interruttore flessibile
Una sorpresa è che una piccola molecola chiamata inositolo esafosfato si annida in una tasca vicino alla fessura catalitica di UBA6. Questo cofattore aiuta a mantenere la scanalatura aperta in una configurazione che accoglie i loop insoliti degli E2 di classe IV, inclusi UBE2Z e BIRC6, rendendo al contempo la stessa regione meno compatibile con gli E2 preferiti da UBA1. Strutture ingrandite di uno stato intermedio, in cui UBA6 e UBE2Z sono legati simultaneamente a due copie di ubiquitina o di FAT10, mostrano come avviene il trasferimento dell’etichetta. Un loop mobile in UBE2Z si sposta di lato come un cancello, esponendo la sua cisteina reattiva in modo che l’etichetta possa essere trasferita. La superficie di contatto può adattarsi: forma un’interfaccia più polare e basata sulle cariche con FAT10 e un’interfaccia più idrofobica, simile a olio, con l’ubiquitina, permettendo allo stesso E2 di gestire due etichette chimicamente distinte.
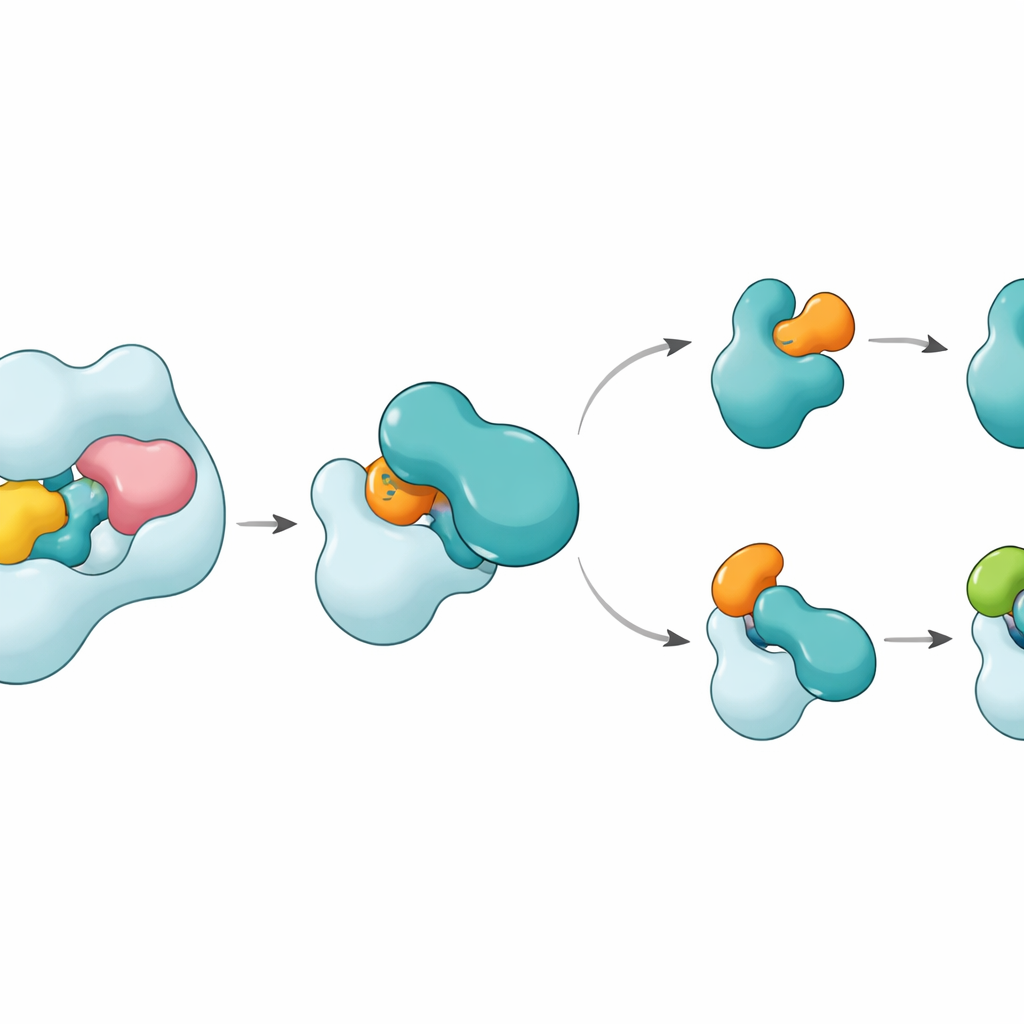
Come questo mantiene l’etichettatura cellulare sulla buona strada
In termini semplici, questo lavoro mostra che UBA6 ha evoluto una culla di riconoscimento specializzata a due parti, rinforzata da un piccolo cofattore, che le consente di lavorare a stretto contatto con un gruppo selezionato di partner E2 e di supportare sia l’etichettatura con ubiquitina sia con FAT10. UBA1, al contrario, utilizza una strategia più tradizionale basata su una singola regione e collabora con un diverso set di E2. Queste soluzioni progettuali distinte creano due rami paralleli ma isolati del sistema di etichettatura nella cellula: uno che gestisce il controllo proteico quotidiano e l’altro che collega lo smaltimento delle proteine ai segnali immunitari. Capire questa logica architetturale non solo chiarisce come le cellule mantengono l’ordine nelle loro reti proteiche, ma potrebbe anche orientare futuri sforzi per mirare rami specifici del sistema di etichettatura in cancro, immunità e altre malattie.
Citazione: Nayak, D., Jia, L., dos Santos Bury, P. et al. Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer. Nat Commun 17, 3302 (2026). https://doi.org/10.1038/s41467-026-69882-3
Parole chiave: ubiquitina, FAT10, UBA6, degradazione delle proteine, cryo-EM