Clear Sky Science · pt
Estruturas por crio‑EM de UBA6 revelam mecanismos da especificidade E1–E2 e da transferência tioéster dual FAT10/ubiquitina
Como as Células Decidem Quais Proteínas Marcar
Dentro de cada célula, um sistema de marcação oculto decide quais proteínas serão enviadas para reciclagem, quais serão remodeladas e quais ajudam a montar uma resposta imune. Duas pequenas etiquetas, ubiquitina e FAT10, participam dessas decisões e precisam ser carregadas em enzimas transportadoras com precisão extrema. Este artigo explora como uma enzima carregadora chave, chamada UBA6, trabalha seletivamente com certos parceiros enzimáticos e lida tanto com ubiquitina quanto com FAT10, revelando princípios de projeto que mantêm o sistema de controle proteico da célula preciso e adaptável.
Duvas Vias Paralelas de Marcação
As células usam etiquetas moleculares para marcar proteínas para destinos diferentes, desde degradação até mudanças de atividade ou localização. A ubiquitina é a etiqueta mais conhecida, empregada em muitos processos celulares, enquanto a FAT10 é mais especializada e fortemente associada a respostas imunes e inflamação. Ambas as etiquetas são ligadas por meio de um revezamento em três etapas envolvendo enzimas E1, E2 e E3. As enzimas E1 realizam o primeiro passo, que consome energia, ativando a etiqueta e transferindo‑a para as E2, que então trabalham com as E3 para anexar a etiqueta às proteínas‑alvo. Os humanos são incomuns por ter duas enzimas E1 para ubiquitina: UBA1, que atende à via clássica da ubiquitina, e UBA6, que não só ativa ubiquitina como também FAT10, formando uma ponte entre o controle proteico cotidiano e a eliminação de proteínas regulada pela imunidade.
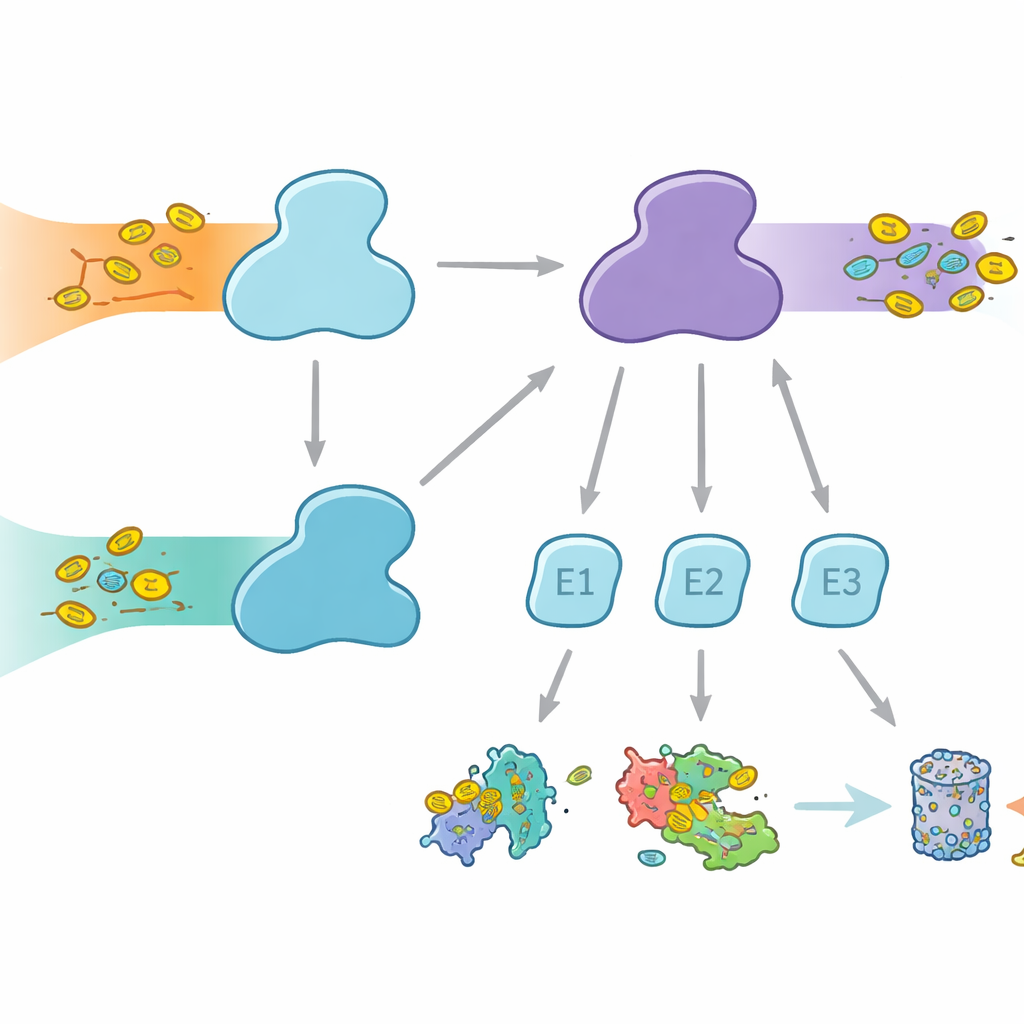
Por Que a Escolha de Parceiros Importa
Embora UBA1 e UBA6 pareçam amplamente similares, elas trabalham com conjuntos em grande parte diferentes de enzimas parceiras E2, criando ramos paralelos do sistema de marcação. Algumas E2s podem funcionar com ambas as E1s, mas outras são parceiras exclusivas. Essa seletividade é vital: se as combinações erradas se formassem, os sinais ligados à FAT10 relacionados à resposta imune poderiam vazar para as vias usuais da ubiquitina, ou vice‑versa, embaralhando decisões celulares. Até agora, não estava claro como UBA6 poderia reconhecer suas E2s preferidas, especialmente um grupo especializado conhecido como E2s classe IV, enquanto UBA1 em grande parte as ignora. Os autores buscaram capturar UBA6 e seus parceiros em ação usando crio‑microscopia eletrônica de alta resolução e experimentos bioquímicos que medem a transferência da etiqueta com grande sensibilidade.
Uma Estratégia de Reconhecimento de Duas Garras
As estruturas revelam que UBA6 reconhece sua parceira E2 chave, chamada UBE2Z, usando um aperto de “duas mãos”. Uma mão é um domínio na extremidade de UBA6 que serve como ponto de ancoragem para as E2s, enquanto a outra é uma região catalítica situada mais próxima do sítio da reação química. Diferentemente de UBA1, que depende majoritariamente da região de ancoragem para selecionar seus parceiros, UBA6 usa ambas as regiões em conjunto para acomodar UBE2Z de modo a alinhar com precisão os dois sítios reativos necessários para a transferência da etiqueta. Segmentos adicionais de loop em UBE2Z, ausentes em muitas outras E2s, encaixam‑se numa ranhura ampliada em UBA6, permitindo um envolvimento firme porém flexível. Se tanto a superfície de ancoragem quanto a cavidade catalítica forem alteradas, UBA6 falha em carregar UBE2Z de maneira eficiente, mostrando que ambas as “mãos” são necessárias para a escolha correta do parceiro.
Um Pequeno Cofator e uma Chave Flexível
Uma surpresa é que uma pequena molécula chamada inositol hexafosfato se aloja numa cavidade próxima à fenda catalítica de UBA6. Esse cofator ajuda a manter a ranhura aberta numa configuração que acolhe os loops incomuns das E2s classe IV, incluindo UBE2Z e BIRC6, ao mesmo tempo que torna a mesma região menos compatível com as E2s favorecidas por UBA1. Estruturas ampliadas de um estado intermediário, onde UBA6 e UBE2Z estão simultaneamente ligados a duas cópias de ubiquitina ou FAT10, mostram como a etiqueta é transferida. Um loop móvel em UBE2Z se afasta como uma cancela, expondo sua cisteína reativa para que a etiqueta possa ser transferida. A superfície de contato pode se adaptar: forma uma interface mais polar e baseada em cargas com FAT10 e uma interface mais hidrofóbica, “oléica”, com ubiquitina, permitindo que a mesma E2 lide com duas etiquetas quimicamente distintas.
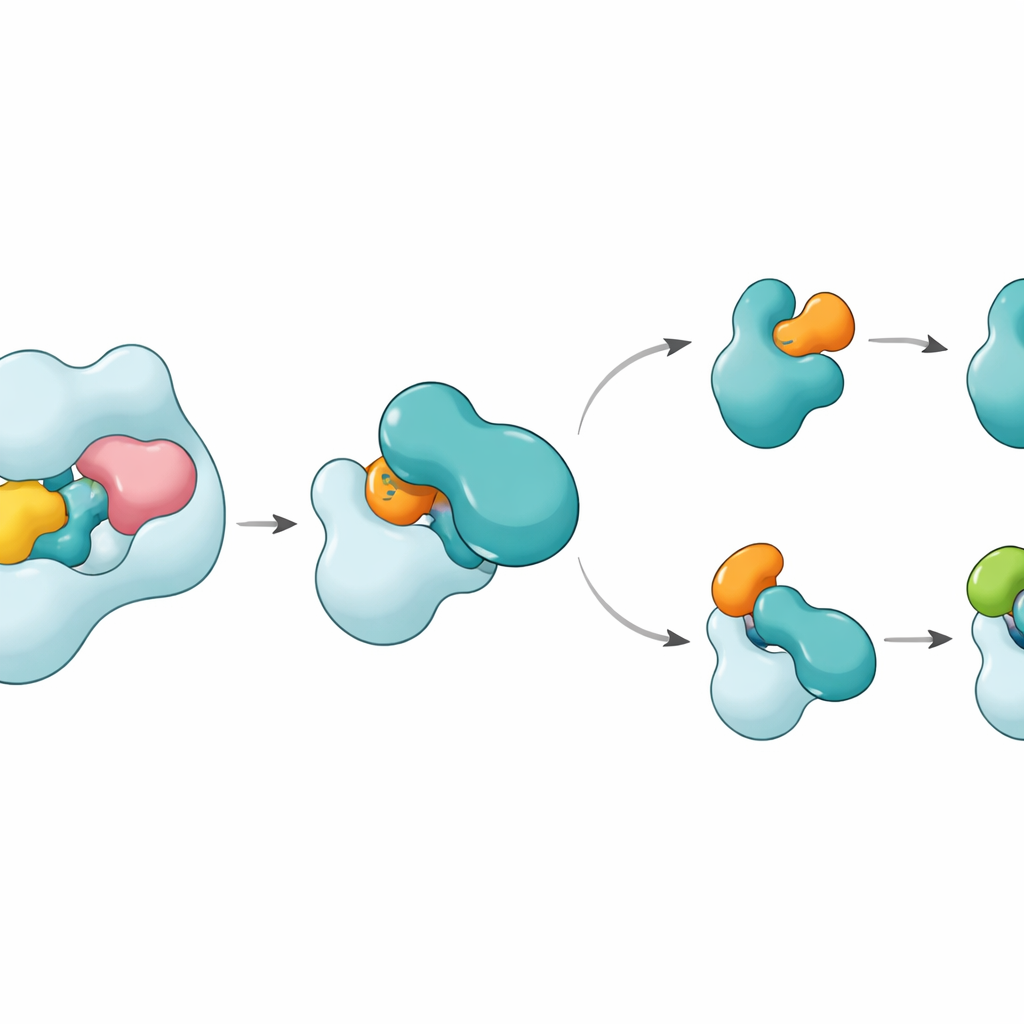
Como Isso Mantém a Marcação Celular no Rumo Certo
Em termos simples, este trabalho mostra que UBA6 evoluiu um berço de reconhecimento especializado em duas partes, reforçado por um pequeno cofator, que lhe permite trabalhar de perto com um grupo seleto de parceiros E2 e suportar tanto a marcação por ubiquitina quanto por FAT10. Em contraste, UBA1 usa uma estratégia mais tradicional, baseada numa única região, e se associa a um conjunto diferente de E2s. Essas soluções de projeto distintas criam dois ramos paralelos, porém isolados, do sistema de marcação dentro da célula: um lidando com o controle proteico cotidiano e outro ligando a eliminação de proteínas a sinais imunes. Compreender essa lógica arquitetural não só esclarece como as células mantêm ordem em suas redes proteicas, mas também pode orientar esforços futuros para direcionar ramos específicos do sistema de marcação em câncer, imunidade e outras doenças.
Citação: Nayak, D., Jia, L., dos Santos Bury, P. et al. Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer. Nat Commun 17, 3302 (2026). https://doi.org/10.1038/s41467-026-69882-3
Palavras-chave: ubiquitina, FAT10, UBA6, degradação de proteínas, crio‑EM