Clear Sky Science · ru
Крио-ЭМ структуры UBA6 раскрывают механизмы специфичности E1–E2 и двойной перенос тиоэстера FAT10/убиквитин
Как клетки решают, какие белки пометить
Внутри каждой клетки скрытая система маркировки решает, какие белки отправить на утилизацию, какие подвергнуть перестройке, а какие задействовать в иммунном ответе. Две крошечные метки — убиквитин и FAT10 — помогают принимать эти решения, и их нужно точно загружать на переносящие ферменты. В этой статье исследуется, как один ключевой загрузчик, называемый UBA6, избирательно взаимодействует с определёнными партнёрными ферментами и одновременно работает с убиквитином и FAT10, что раскрывает принципы организации, сохраняющие точность и адаптивность клеточной системы контроля белков.
Две параллельные магистрали маркировки
Клетки используют молекулярные метки, чтобы помечать белки для разных судьб — от деградации до изменения активности или локализации. Убиквитин — наиболее известная метка, задействованная во множестве клеточных процессов, тогда как FAT10 более специализирован и тесно связан с иммунными ответами и воспалением. Обе метки присоединяются через трёхступенчатую эстафету с участием ферментов E1, E2 и E3. Ферменты E1 выполняют первый, энергозатратный этап, активируя метку и передавая её E2, которые затем в сотрудничестве с E3 присоединяют метку к целевым белкам. У людей необычная особенность — два фермента E1 для убиквитина: UBA1, обслуживающий классический путь убиквитинирования, и UBA6, который активирует не только убиквитин, но и FAT10, создавая мост между повседневным контролем белков и иммунно‑регулируемой утилизацией белков.
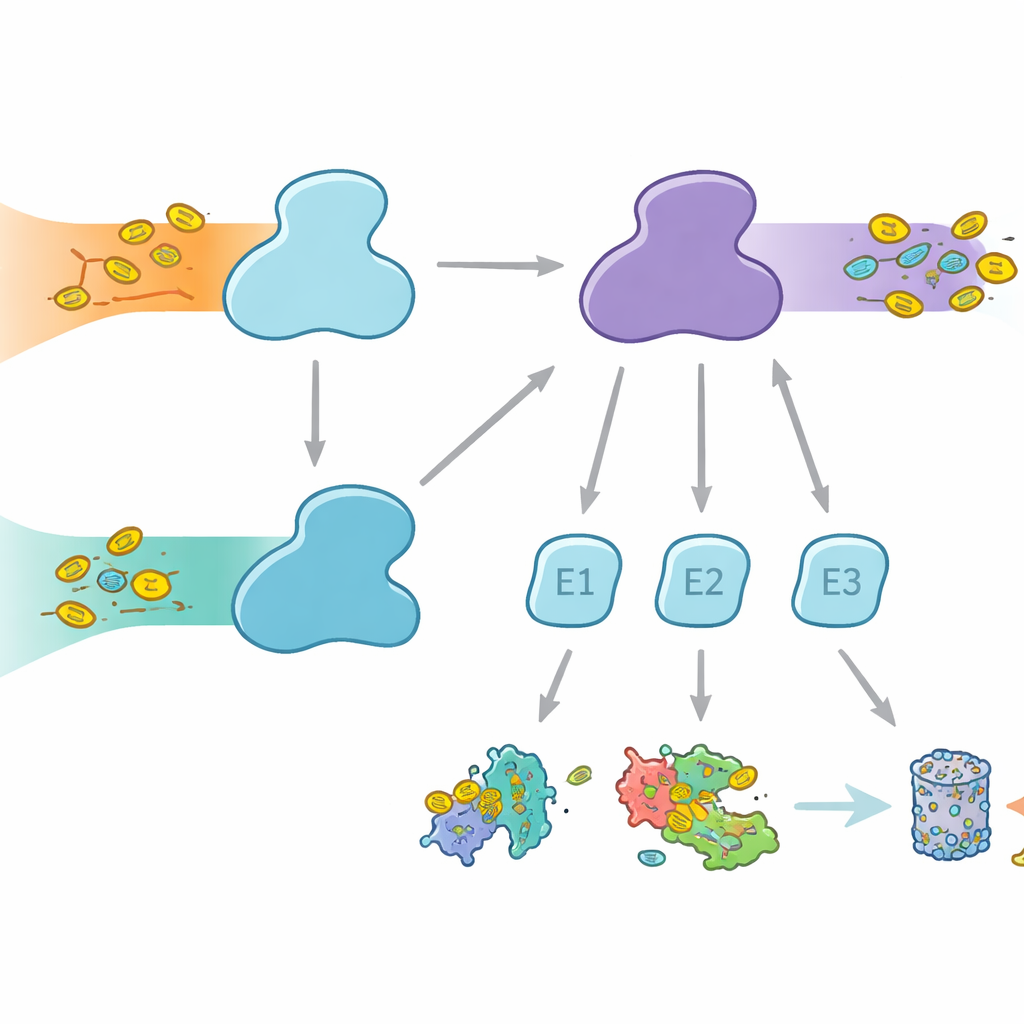
Почему выбор партнёра важен
Хотя UBA1 и UBA6 в целом схожи по виду, они работают преимущественно с разными наборами партнёрных E2‑ферментов, создавая параллельные ветви системы маркировки. Некоторые E2 могут взаимодействовать с обоими E1, но другие являются эксклюзивными партнёрами. Такая селективность жизненно важна: если бы формировались неверные сочетания, сигналы FAT10, связанные с иммунитетом, могли бы «протекать» в обычные пути убиквитинирования или наоборот, нарушая клеточные решения. До сих пор было непонятно, как UBA6 распознаёт своих предпочтительных E2, особенно специализированную группу, известную как класс IV E2, в то время как UBA1 их в основном игнорирует. Авторы поставили задачу запечатлеть UBA6 и его партнёров в действии с помощью высокоразрешающей крио‑электронной микроскопии и биохимических экспериментов, измеряющих передачу меток с высокой чувствительностью.
Стратегия распознавания с двумя «захватами»
Структуры показывают, что UBA6 распознаёт своего ключевого партнёра E2, называемого UBE2Z, с помощью «двуручного» захвата. Одна «рука» — это домен на конце UBA6, служащий докинг‑площадкой для E2, а другая — каталитическая область, расположенная ближе к месту химической реакции. В отличие от UBA1, который в основном полагается на докинг‑регион при выборе партнёров, UBA6 использует обе области вместе, чтобы аккуратно зажать UBE2Z и точно выровнять два реактивных центра, необходимые для переноса метки. Дополнительные сегменты петель на UBE2Z, отсутствующие у многих других E2, входят в расширенную выемку в UBA6, обеспечивая плотное, но гибкое взаимодействие. Если изменить либо докинг‑поверхность, либо каталитическую впадину, UBA6 неэффективно загружает UBE2Z, что показывает, что обе «руки» требуются для правильного выбора партнёра.
Малый кофактор и гибкий переключатель
Одной из неожиданностей оказалось присутствие крошечной молекулы — инозитолгексаксифосфата — в кармане рядом с каталитической впадиной UBA6. Этот кофактор помогает удерживать выемку в конфигурации, благоприятной для приёма необычных петель класс IV E2, включая UBE2Z и BIRC6, одновременно делая ту же область менее совместимой с E2, предпочитаемыми UBA1. Увеличенные в деталях структуры промежуточного состояния, в котором UBA6 и UBE2Z одновременно связаны с двумя копиями либо убиквитина, либо FAT10, показывают, как происходит передача метки. Подвижная петля в UBE2Z отодвигается как створка, обнажая реактивную цистеиновую группу, чтобы метка могла быть передана. Контактная поверхность может адаптироваться: с FAT10 она образует более полярный, заряд‑зависимый интерфейс, а с убиквитином — более гидрофобный, «маслянистый» интерфейс, что позволяет одному и тому же E2 работать с двумя химически различными метками.
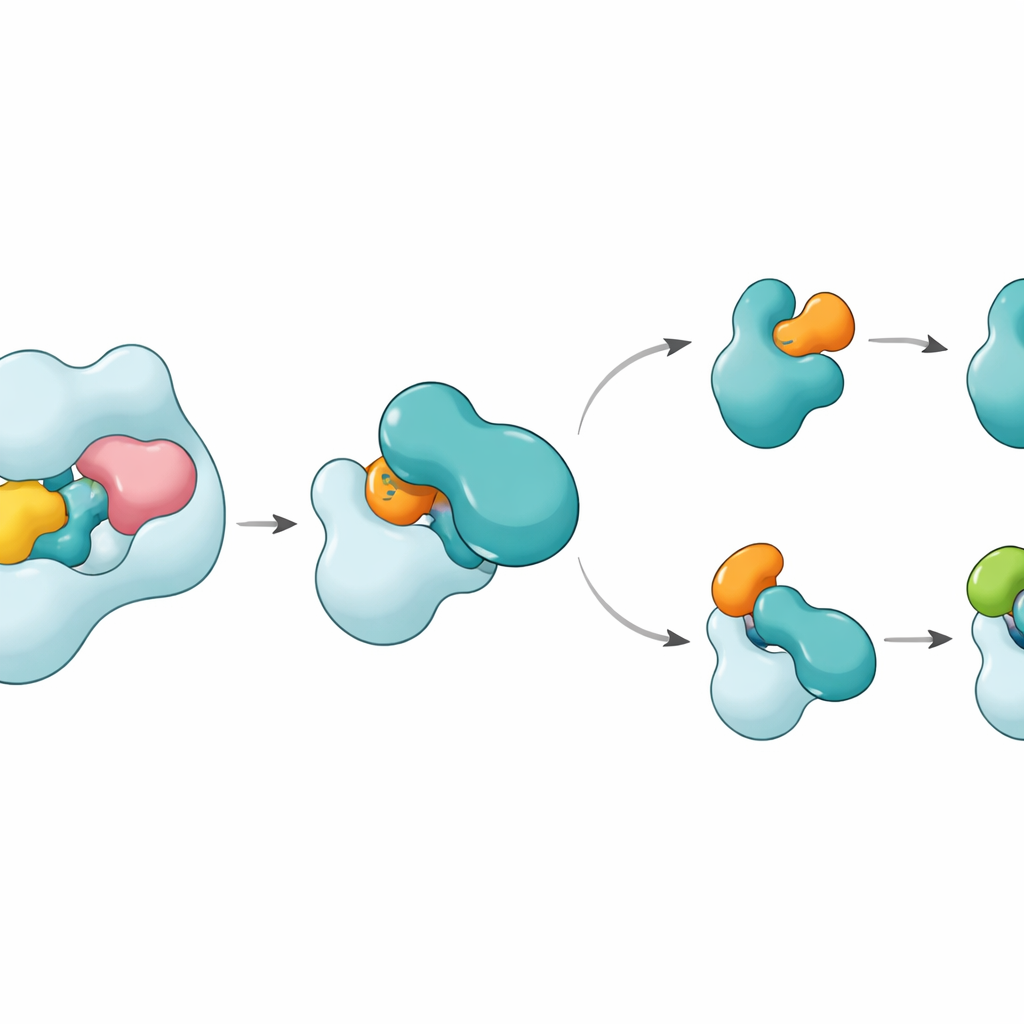
Как это сохраняет порядок в маркировке клеток
Проще говоря, эта работа показывает, что UBA6 развил специализированную, двухчастную «ракушку» распознавания, усиленную малым кофактором, что позволяет ему тесно сотрудничать с избранной группой партнёрных E2 и поддерживать как убиквитин‑, так и FAT10‑маркировку. В отличие от него UBA1 использует более традиционный подход с одной областью и сотрудничает с другим набором E2. Эти разные архитектурные решения создают внутри клетки две параллельные, но изолированные ветви маркировки: одну — для повседневного контроля белков, другую — связывающую утилизацию белков с иммунными сигналами. Понимание этой архитектурной логики не только разъясняет, как клетки поддерживают порядок в сетях белков, но и может помочь в будущих усилиях по целенаправленной модификации отдельных ветвей системы маркировки в онкологии, иммунологии и других заболеваниях.
Цитирование: Nayak, D., Jia, L., dos Santos Bury, P. et al. Cryo-EM structures of UBA6 reveal mechanisms of E1–E2 specificity and dual FAT10/ubiquitin thioester transfer. Nat Commun 17, 3302 (2026). https://doi.org/10.1038/s41467-026-69882-3
Ключевые слова: убиквитин, FAT10, UBA6, деградация белков, крио-ЭМ