Clear Sky Science · ru
Единая модель кодирования скорости для беговых и отходных реакций в раздельных нейронных цепях
Как мозг задаёт темп побега
Когда животное решает, броситься вперёд, затаиться или осторожно отступить, его мозг должен не только выбрать действие, но и определить, с какой скоростью это действие будет выполняться. Исследование на мышах раскрывает, как крошечный участок глубоко в мозге вычисляет скорость движения и направляет этот общий сигнал скорости в совсем разные оборонительные реакции. Работа даёт представление о том, как мозг превращает смутные ощущения угрозы в точно настроенное движение и почему одна и та же опасность иногда заставляет нас мчаться вперёд, а иногда — отступать или останавливаться.
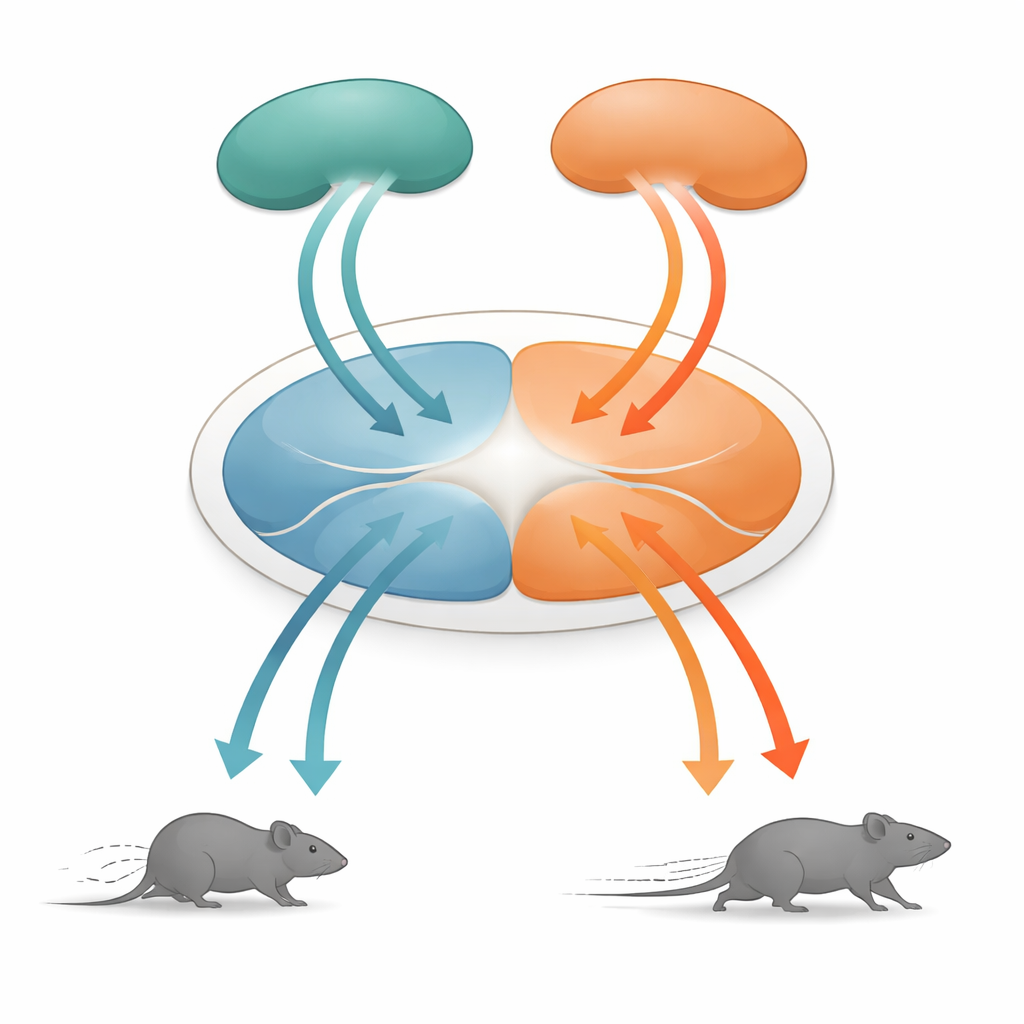
Центральный узел для выживательных решений
В центре этой истории — дорсальная периакадуктальная серая субстанция (dPAG), небольшая структура в среднем мозге, давно известная как критическая для оборонительных поведений. Она получает информацию от многих высших областей мозга и преобразует эти сигналы в конкретные движения. Исследователи сосредоточились на двух основных источниках входов: височной ассоциативной коре (TeA), которая помогает связывать сенсорные события в контексте, и верхней затылочной (колликулюс) — superior colliculus (SC), структуре, быстро обнаруживающей нависавшие визуальные угрозы. Обе области проецируют в dPAG, но до сих пор было неясно, как их активность превращается в детализованные моторные команды, такие как скорость бега.
Нейроны, предсказывающие, как быстро вы побежите
С помощью тонких записей отдельных нейронов у бодрствующих мышей с зафиксированной головой, бегавших по вращающейся платформе, команда сначала выделила клетки в TeA, частота спайков которых росла и падала в соответствии со скоростью бега животного. Эти «связанные с бегом» нейроны не реагировали напрямую на вспышки света, звуки или потоки воздуха; вместо этого их активность нарастала за одну–три секунды до начала бега и продолжала отражать скорость во время движения. При построении графика частоты спайков против скорости бега выяснилось, что зависимость подчинялась насыщающейся кривой: при увеличении активности скорость сначала быстро возрастала, а затем выравнивалась на максимуме, что соответствует биологическому пределу — животные не могут ускоряться бесконечно.
Общий код скорости в более глубоком слое мозга
Аналогичный тип эксперимента в dPAG выявил параллельную картину. Нейроны там также усиливали свою активность до и во время бега, а их соотношение активности и скорости следовало той же математической кривой, что и в TeA. Однако нейроны dPAG были более «эффективными»: при данной смене активности они вызывали большие изменения в скорости бега, и их влияние начинало сказываться ближе ко времени начала движения. Стимулируя специфические наборы нейронов TeA и dPAG светом с разной частотой, исследователи причинно контролировали скорость бега в соответствии с той же кривой. Это показало, что отношение — не просто корреляция, а подлинное правило кодирования скорости, общее для разных уровней иерархии движения.
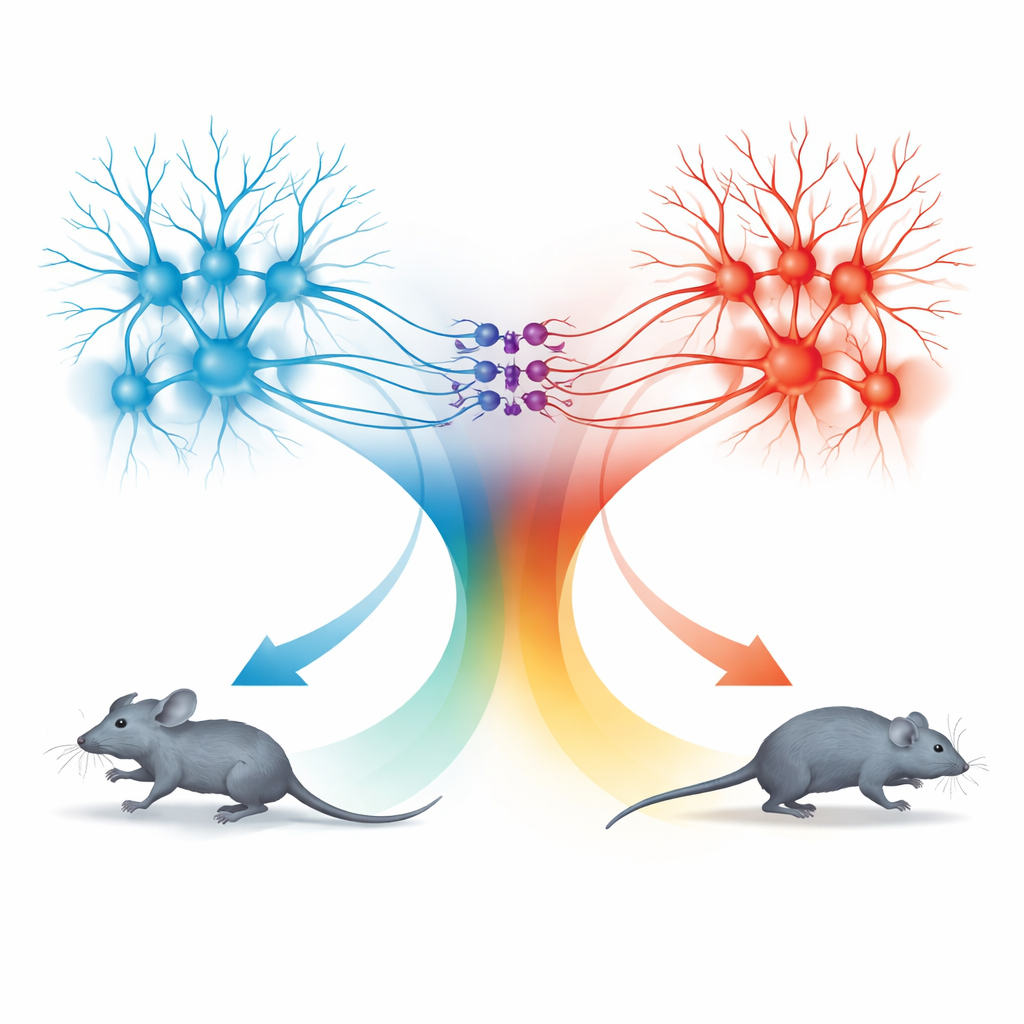
Два блока движения: бег вперёд и отступление
Сюрприз обнаружился при исследовании пути от верхнего колликулюса в dPAG. Нейроны в этом пути также следовали тому же правилу скорости, но поведение, которое они вызывали, было иным. При активации этих соединённых со SC клеток низкими частотами света мыши прекращали движение; при более высоких частотах они отступали назад; а по окончании стимуляции у них часто возникал короткий всплеск быстрого бега вперёд, называемый реактивным (rebound) бегом. Детальная анатомия показала, что сигналы из TeA и SC поступали в основном в разные популяции нейронов dPAG. Возбудающие клетки, связанные с TeA, формировали «блок бега», ответственный за прямолинейный побег, тогда как возбуждающие клетки, связанные с SC, образовывали «блок отступления». Небольшой набор ингибирующих соматостатин-позитивных (SOM) нейронов в dPAG соединял эти блоки в одном направлении: они активировались блоком отступления и в свою очередь подавляли блок бега.
Простая схема для множества оборонительных состояний
Комбинируя селективную активацию и выключение, команда отобразила, как эти три типа клеток взаимодействуют, порождая четыре различные поведения: бег, отступление, остановку и реактивный бег. Активация только клеток, связанных с TeA, вызывала вперёд бег, чья скорость следовала общей кодировочной кривой. Активация клеток, связанных с SC, включала блок отступления и SOM-нейроны, подавляя блок бега и вызывая либо остановку, либо движение назад в зависимости от силы стимуляции. Когда стимуляция прекращалась, внезапное снятие SOM-зависимого торможения позволяло блоку бега кратковременно «отскочить», давая всплеск вперёд. Таким образом dPAG использует единый количественный код скорости, но прокладывает его через разные микросхемы, чтобы генерировать разные оборонительные варианты.
Что это означает для понимания выбора действий
Для неспециалиста ключевая мысль такова: мозг разделяет две задачи — насколько сильным должно быть движение и какое именно движение выполнять. В этой модели на мышах общее математическое правило преобразует нейронную активность в скорость движения, а специализированные «блоки поведения» в dPAG решают, превратится ли эта скорость в бег вперёд или в отступление. Небольшой ингибирующий мост позволяет системе быстро переключаться между этими состояниями, даже вызывая реактивный рывок после периода вынужденной неподвижности. Эта работа является наглядным примером того, как компактные нейронные цепи гибко оркестрируют сложные выживательные поведения, используя простые, повторно применимые правила кодирования.
Цитирование: Chen, J., Li, H., Lian, N. et al. A shared speed encoding model for running and backing away behaviours in segregated neural circuits. Nat Commun 17, 4119 (2026). https://doi.org/10.1038/s41467-026-70755-y
Ключевые слова: оборонительное поведение, скорость движения, стволовая части мозга, реакции бегства, нейронаука на мышах