Clear Sky Science · es
Un modelo compartido de codificación de la velocidad para comportamientos de carrera y retroceso en circuitos neuronales segregados
Cómo el cerebro fija el ritmo del escape
Cuando un animal decide si lanzarse hacia delante, quedarse inmóvil o retroceder con precaución, su cerebro no solo debe elegir una acción sino también determinar la velocidad a la que se ejecuta. Este estudio en ratones revela cómo una pequeña región profunda del cerebro calcula la velocidad del movimiento y dirige esa señal de velocidad compartida hacia respuestas defensivas muy distintas. El trabajo ofrece una ventana sobre cómo el cerebro convierte sensaciones vagas de amenaza en movimiento finamente ajustado, y por qué el mismo peligro a veces nos hace precipitar hacia delante y otras veces retroceder o detenernos.
Un núcleo central para decisiones de supervivencia
En el centro de esta historia está la sustancia gris periacueductal dorsal, o dPAG, una pequeña estructura del mesencéfalo conocida desde hace tiempo por su papel crucial en comportamientos defensivos. Recibe información de muchas zonas cerebrales superiores y convierte esas señales en movimientos concretos. Los investigadores se centraron en dos fuentes de entrada principales: la corteza de asociación temporal (TeA), que ayuda a vincular eventos sensoriales con el contexto, y el colículo superior (SC), una estructura que detecta con rapidez amenazas visuales en descenso. Ambas regiones envían proyecciones a la dPAG, pero hasta ahora no quedaba claro cómo su actividad se transforma en comandos motores detallados como la velocidad de carrera.
Neuronas que predicen la rapidez con la que correrás
Con registros delicados de neuronas individuales en ratones despiertos con la cabeza fijada corriendo sobre una plataforma giratoria, el equipo identificó primero células en la TeA cuyos ritmos de disparo aumentaban y disminuían con la velocidad de carrera del animal. Estas neuronas “relacionadas con la carrera” no respondían directamente a destellos, sonidos o soplos de aire; en cambio, su actividad aumentaba entre uno y tres segundos antes de que el animal comenzara a correr y continuaba reflejando su velocidad durante la carrera. Al trazar la tasa de disparo frente a la velocidad de carrera, los científicos encontraron que la relación seguía una curva de saturación: a medida que la actividad aumentaba, la velocidad se elevaba rápido al principio y luego se estabilizaba en un máximo, concordando con el límite biológico de que los animales no pueden acelerar indefinidamente.
Un código de velocidad compartido en una capa cerebral más profunda
El mismo tipo de experimento en la dPAG reveló un patrón paralelo. Allí las neuronas también aumentaban su disparo antes y durante la carrera, y la relación entre disparo y velocidad seguía la misma curva matemática que en la TeA. Sin embargo, las neuronas de la dPAG eran más “eficientes”: para un mismo cambio en la actividad neuronal producían mayores cambios en la velocidad de carrera, y su influencia se activaba más cerca del inicio del movimiento. Estimulando conjuntos específicos de neuronas de la TeA y la dPAG con luz a diferentes frecuencias, los investigadores pudieron ajustar causalmente la velocidad de carrera hacia arriba o hacia abajo según esa misma curva. Esto mostró que la relación no es solo una correlación, sino una regla genuina de codificación de la velocidad compartida a diferentes niveles de la jerarquía del movimiento.
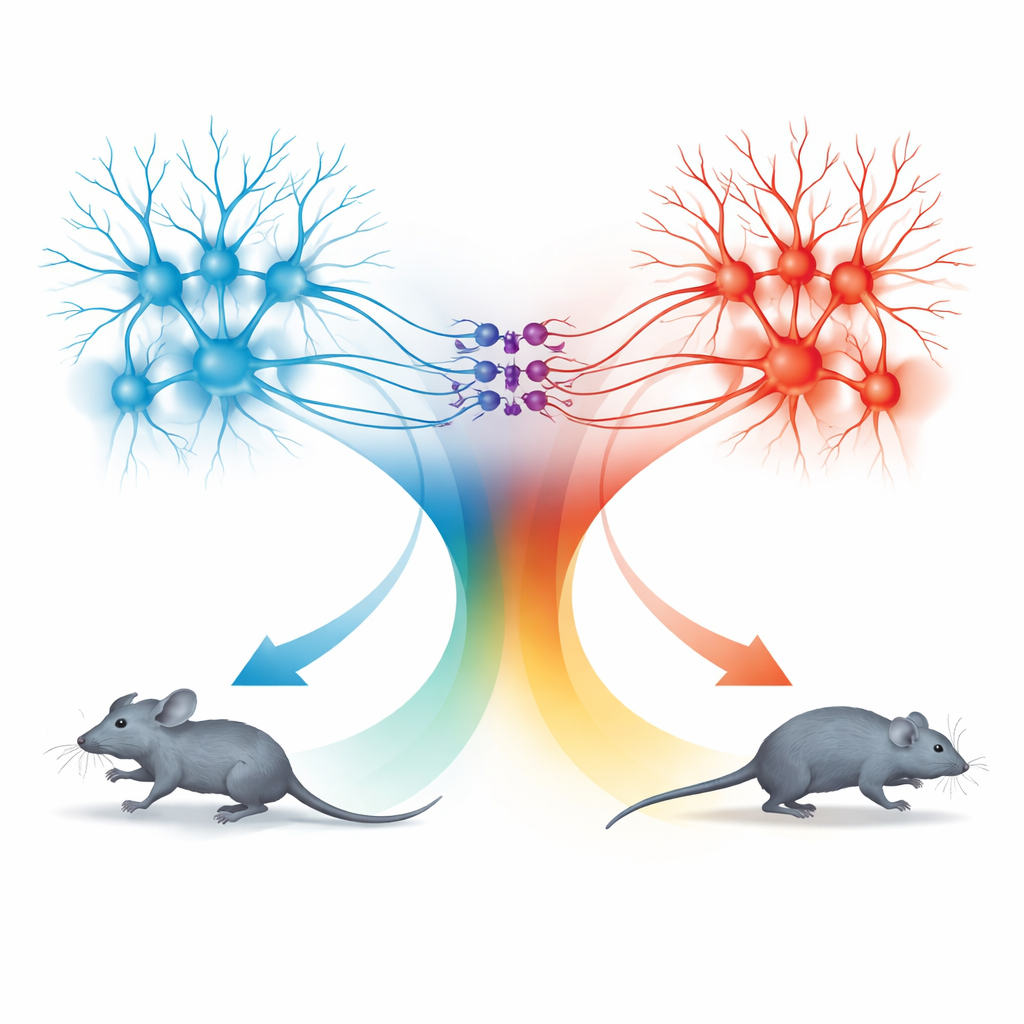
Dos unidades de movimiento: correr hacia delante y retroceder
La sorpresa vino al examinar la vía desde el colículo superior hacia la dPAG. Las neuronas en esta vía también seguían la misma regla de velocidad, pero los comportamientos que desencadenaban eran distintos. Cuando estas células conectadas al SC se activaban con frecuencias lumínicas bajas, los ratones dejaban de moverse; a frecuencias mayores retrocedían; y cuando la estimulación terminaba, a menudo estallaban en un breve período de carrera rápida hacia delante, llamada carrera de rebote. Trazados detallados mostraron que las señales de TeA y SC llegaban a poblaciones en gran parte separadas de neuronas dPAG. Las células excitadoras conectadas a la TeA formaban una “unidad de carrera” que impulsaba la huida hacia delante, mientras que las células excitadoras conectadas al SC formaban una “unidad de retroceso”. Un pequeño conjunto de neuronas inhibitorias de somatostatina (SOM) en la dPAG conectaba estas unidades en una sola dirección: eran activadas por la unidad de retroceso y, a su vez, suprimían la unidad de carrera.
Un circuito sencillo para muchos estados defensivos
Combinando activación y silenciamiento selectivo, el equipo mapeó cómo estos tres tipos celulares se combinan para producir cuatro comportamientos distintos: correr, retroceder, detenerse y carrera de rebote. Activar solo las células conectadas a la TeA producía carrera hacia delante cuya velocidad seguía la curva de codificación compartida. Activar las células conectadas al SC ponía en marcha la unidad de retroceso y las neuronas SOM, apagando la unidad de carrera y provocando detención o movimiento hacia atrás según la intensidad de la estimulación. Cuando la estimulación cesaba, el levantamiento súbito de la inhibición mediada por SOM permitía que la unidad de carrera rebotase brevemente, produciendo un estallido de escape hacia delante. De este modo, la dPAG utiliza un único código cuantitativo de velocidad pero lo enruta a través de microcircuitos distintos para generar diferentes elecciones defensivas.
Qué implica esto para entender la elección de acciones
Para un público no especializado, el mensaje clave es que el cerebro separa dos problemas: cuán potente debe ser un movimiento y qué movimiento ejecutar. En este modelo de ratón, una regla matemática compartida transforma el disparo neuronal en velocidad de movimiento, mientras que unidades de comportamiento especializadas en la dPAG deciden si esa velocidad se traduce en correr hacia delante o en retroceder. Un pequeño puente inhibitorio permite entonces al sistema cambiar rápidamente entre estos estados, incluso produciendo un impulso de huida tras un periodo de inmovilidad forzada. Este trabajo ofrece un ejemplo concreto de cómo circuitos neuronales compactos pueden orquestar de forma flexible comportamientos complejos de supervivencia usando reglas de codificación simples y reutilizables.
Cita: Chen, J., Li, H., Lian, N. et al. A shared speed encoding model for running and backing away behaviours in segregated neural circuits. Nat Commun 17, 4119 (2026). https://doi.org/10.1038/s41467-026-70755-y
Palabras clave: comportamiento defensivo, velocidad de movimiento, circuitos del mesencéfalo, respuestas de escape, neurociencia en ratones