Clear Sky Science · pt
Um modelo compartilhado de codificação de velocidade para comportamentos de corrida e recuo em circuitos neurais segregados
Como o cérebro define o ritmo da fuga
Quando um animal decide se deve disparar para frente, congelar ou recuar cautelosamente, seu cérebro precisa não apenas escolher uma ação, mas também determinar a velocidade com que essa ação ocorrerá. Este estudo em camundongos revela como uma pequena região profunda do cérebro computa a velocidade do movimento e encaminha esse sinal de velocidade compartilhado para respostas defensivas muito diferentes. O trabalho oferece uma janela sobre como o cérebro transforma sensações vagas de ameaça em movimento precisamente ajustado, e por que o mesmo perigo às vezes nos leva a avançar apressadamente e outras vezes a recuar ou parar.
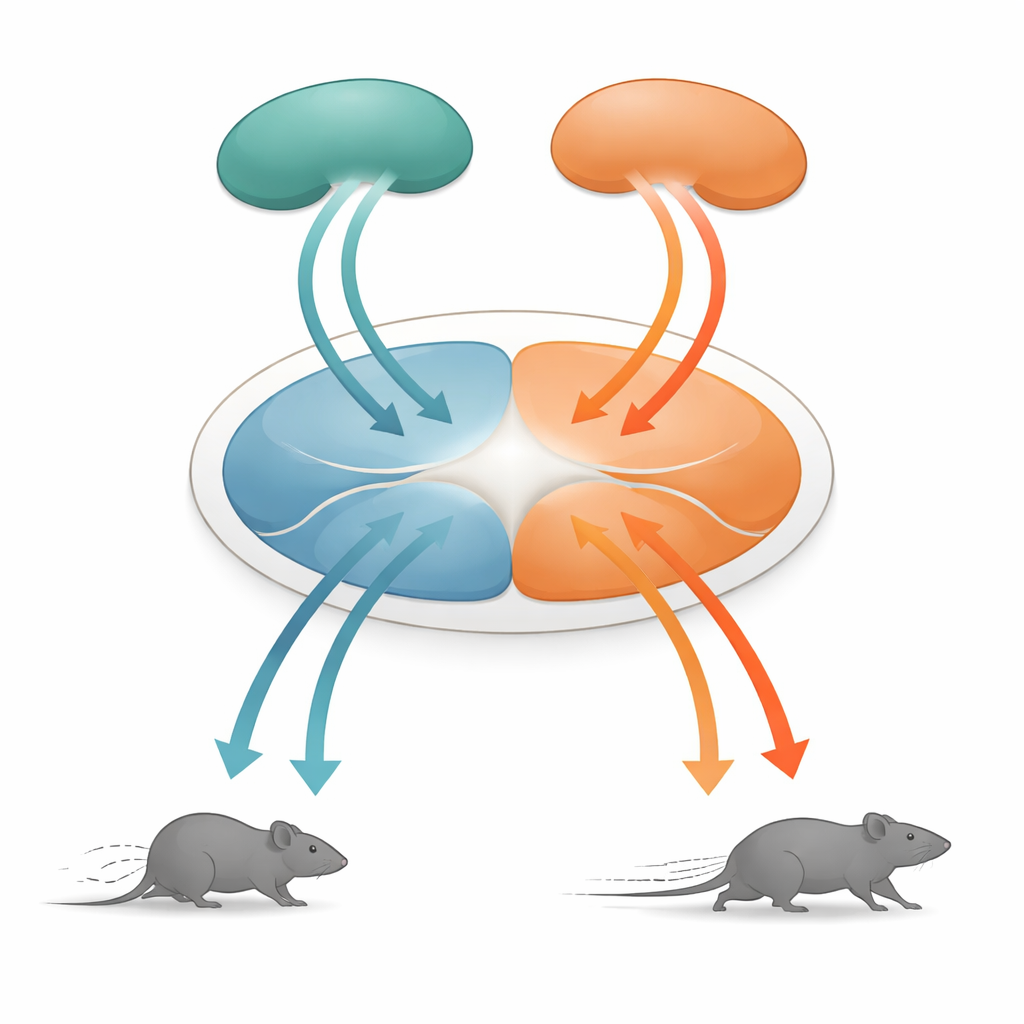
Um centro central para escolhas de sobrevivência
No centro desta história está a substância cinzenta periacuedutal dorsal, ou dPAG, uma pequena estrutura no mesencéfalo conhecida há muito tempo por ser crucial para comportamentos defensivos. Ela recebe informações de muitas áreas cerebrais superiores e converte esses sinais em movimentos concretos. Os pesquisadores se concentraram em duas fontes de entrada principais: o córtex de associação temporal (TeA), que ajuda a ligar eventos sensoriais ao contexto, e o colículo superior (SC), uma estrutura que detecta rapidamente ameaças visuais de aproximação. Ambas as regiões enviam projeções para a dPAG, mas até agora não estava claro como a atividade delas se transforma em comandos motores detalhados como a velocidade de corrida.
Neurônios que predizem quão rápido você correrá
Usando registros delicados de neurônios individuais em camundongos acordados e com a cabeça fixada correndo sobre um prato giratório, a equipe primeiro acompanhou células no TeA cujas taxas de disparo aumentavam e diminuíam com a velocidade de corrida do animal. Esses neurônios “relacionados à corrida” não respondiam diretamente a flashes, sons ou jatos de ar; em vez disso, seu disparo aumentava de um a três segundos antes do início da corrida e continuava a refletir a velocidade durante a corrida. Quando os cientistas traçaram a taxa de disparo em relação à velocidade de corrida, descobriram que a relação seguia uma curva de saturação: à medida que o disparo aumentava, a velocidade subia rapidamente no início e depois se estabilizava em um máximo, compatível com o limite biológico de que os animais não podem acelerar indefinidamente.
Um código de velocidade compartilhado em uma camada cerebral mais profunda
O mesmo tipo de experimento na dPAG revelou um padrão paralelo. Neurônios ali também aumentavam seu disparo antes e durante a corrida, e a relação disparo–velocidade seguia a mesma curva matemática observada no TeA. No entanto, os neurônios da dPAG eram mais “eficientes”: para uma dada mudança no disparo, eles produziam alterações maiores na velocidade de corrida, e sua influência surgia mais próxima do início do movimento. Ao estimular conjuntos específicos de neurônios do TeA e da dPAG com luz em diferentes frequências, os pesquisadores puderam ajustar causalmente a velocidade de corrida para cima e para baixo em conformidade com essa mesma curva. Isso mostrou que a relação não é apenas uma correlação, mas uma regra genuína de codificação de velocidade compartilhada entre níveis da hierarquia de movimento.
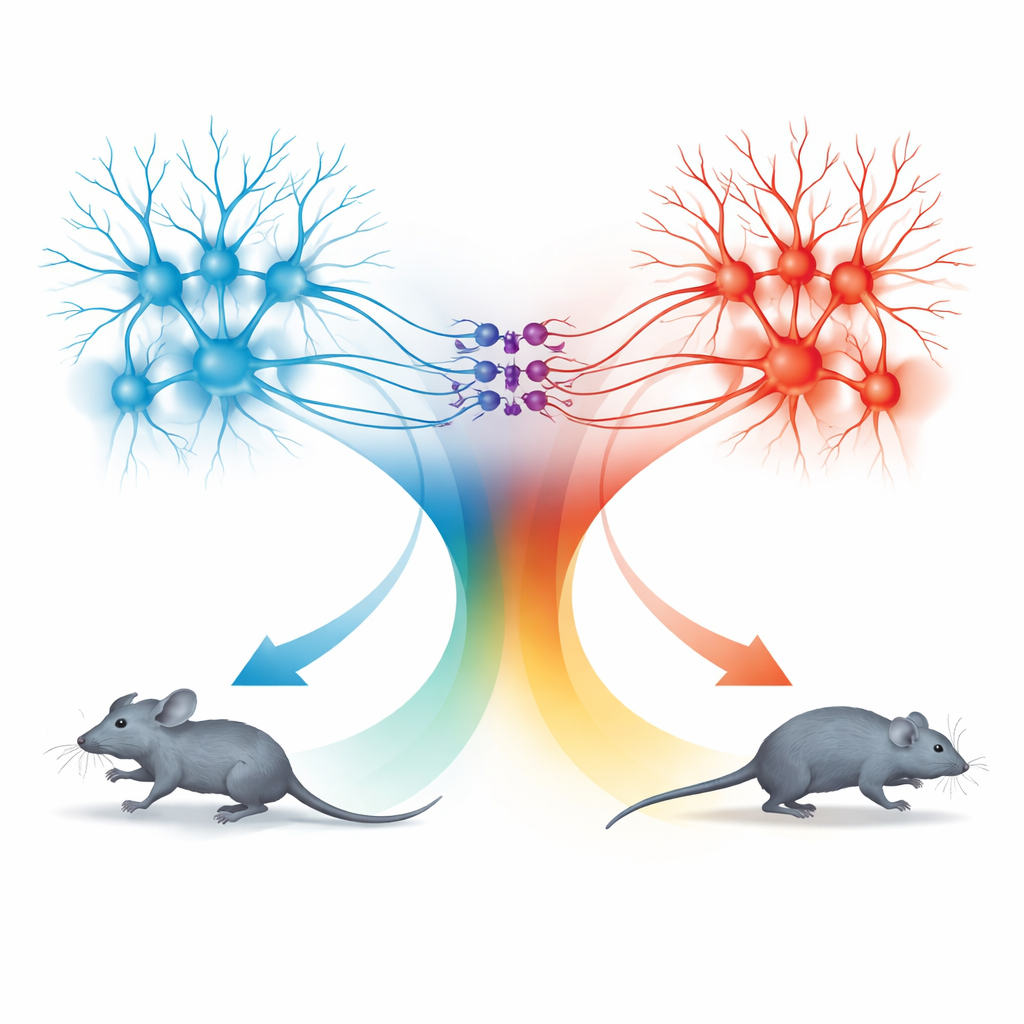
Dupla de unidades de movimento: corrida para frente e recuo
A surpresa veio ao examinar a via do colículo superior para a dPAG. Neurônios nessa rota também seguiam a mesma regra de velocidade, mas os comportamentos que provocavam eram diferentes. Quando essas células conectadas ao SC eram ativadas com baixas frequências de luz, os camundongos paravam de se mover; em frequências mais altas, recuavam; e quando a estimulação terminava, frequentemente entravam em um breve período de corrida rápida para frente, chamado corrida de rebote. Traçagens detalhadas mostraram que os sinais do TeA e do SC alcançavam populações de neurônios da dPAG em grande parte separadas. Células excitatórias ligadas ao TeA formaram uma “unidade de corrida” que impulsionava fugas diretas para frente, enquanto células excitatórias ligadas ao SC formaram uma “unidade de recuo”. Um pequeno conjunto de neurônios inibitórios somatostatina (SOM) na dPAG conectava essas unidades em uma direção: eles eram ativados pela unidade de recuo e, por sua vez, suprimiam a unidade de corrida.
Um circuito simples para muitos estados defensivos
Combinando ativação e silenciamento seletivos, a equipe mapeou como esses três tipos celulares se combinam para gerar quatro comportamentos distintos: correr, recuar, parar e corrida de rebote. Ativar apenas as células ligadas ao TeA produziu corrida para frente cuja velocidade seguia a curva de codificação compartilhada. Ativar as células ligadas ao SC ligava a unidade de recuo e os neurônios SOM, desligando a unidade de corrida e causando ou a parada ou o movimento para trás, dependendo da intensidade da estimulação. Quando a estimulação cessava, o levantamento súbito da inibição mediada por SOM permitia que a unidade de corrida rebentasse brevemente, produzindo um surto de fuga para frente. Dessa forma, a dPAG usa um único código quantitativo de velocidade, mas o encaminha por microcircuitos diferentes para gerar escolhas defensivas distintas.
O que isso significa para entender a escolha de ações
Para um leitor não especialista, a mensagem principal é que o cérebro separa dois problemas: quão forte um movimento deve ser e qual movimento executar. Neste modelo de camundongo, uma regra matemática compartilhada transforma o disparo neural em velocidade de movimento, enquanto “unidades de comportamento” especializadas na dPAG decidem se essa velocidade se torna corrida para frente ou recuo. Uma pequena ponte inibitória então permite que o sistema mude rapidamente entre esses estados, chegando até a produzir um arrancada de rebote após um período de imobilidade forçada. Este trabalho oferece um exemplo concreto de como circuitos neurais compactos podem orquestrar de forma flexível comportamentos complexos de sobrevivência usando regras de codificação simples e reutilizáveis.
Citação: Chen, J., Li, H., Lian, N. et al. A shared speed encoding model for running and backing away behaviours in segregated neural circuits. Nat Commun 17, 4119 (2026). https://doi.org/10.1038/s41467-026-70755-y
Palavras-chave: comportamento defensivo, velocidade de movimento, circuitos do mesencéfalo, respostas de fuga, neurociência em camundongos