Clear Sky Science · nl
Een gedeeld snelheidscodemodel voor vooruit rennen en achteruit terugtrekken in gescheiden neurale circuits
Hoe de hersenen het tempo van de vlucht bepalen
Wanneer een dier beslist om vooruit weg te rennen, te bevriezen of voorzichtig achteruit te wijken, moet zijn brein niet alleen een actie kiezen maar ook bepalen hoe snel die actie verloopt. Deze studie in muizen onthult hoe een klein gebied diep in de hersenen bewegingssnelheid berekent en dat gedeelde snelheidsignaal naar zeer verschillende verdedigingsreacties leidt. Het werk biedt inzicht in hoe het brein vage gevoelens van dreiging omzet in precies afgestemde bewegingen, en waarom hetzelfde gevaar ons soms juist naar voren doet stormen en andere keren doet terugtrekken of stoppen.
Een centraal knooppunt voor overlevingskeuzes
Centraal in dit verhaal staat de dorsale periaqueductale grijze stof, of dPAG, een kleine structuur in de middenhersenen die al lang bekendstaat als cruciaal voor verdedigingsgedrag. Hij ontvangt informatie van veel hogere hersengebieden en zet die signalen om in concrete bewegingen. De onderzoekers richtten zich op twee belangrijke inputbronnen: de temporaalkorticale associatiezone (TeA), die helpt om zintuiglijke gebeurtenissen in context te plaatsen, en de superior colliculus (SC), een structuur die snel dreigende visuele prikkels detecteert. Beide regio’s zenden projecties naar de dPAG, maar tot nu toe was onduidelijk hoe hun activiteit wordt omgezet in gedetailleerde motorcommando’s zoals loopsnelheid.
Neuronenen die voorspellen hoe snel je zult rennen
Met fijne opnamen van individuele neuronen in wakker, met het hoofd gefixeerde muizen die op een draaischijf renden, volgde het team eerst cellen in de TeA waarvan de vuurfrekenties omhoog en omlaag gingen met de loopsnelheid van het dier. Deze "loop-gerelateerde" neuronen reageerden niet direct op flitsen, geluiden of luchtstoten; in plaats daarvan bouwde hun activiteit één tot drie seconden op voordat het dier begon te rennen, en bleef die tijdens het rennen de snelheid weerspiegelen. Wanneer de wetenschappers vuurfrekentie uitzetten tegen loopsnelheid, bleek de relatie een verzadigende curve te volgen: naarmate de vuurfrekentie toenam, steeg de snelheid aanvankelijk snel en vlakte vervolgens af naar een maximum, overeenkomend met de biologische limiet dat dieren niet onbeperkt kunnen versnellen.
Een gedeelde snelheidscode in een dieper hersenlaag
Een vergelijkbaar experiment in de dPAG liet een parallel patroon zien. Neuronen daar verhoogden hun activiteit ook vóór en tijdens het rennen, en hun relatie tussen vuurfrekentie en snelheid volgde dezelfde wiskundige curve als in de TeA. dPAG-neuronen waren echter efficiënter: voor eenzelfde verandering in vuurfrekentie veroorzaakten ze grotere veranderingen in loopsnelheid, en hun invloed trad dichter bij het begin van de beweging in werking. Door specifieke sets TeA- en dPAG-neuronen met licht bij verschillende frequenties te stimuleren, konden de onderzoekers causaal de loopsnelheid omhoog en omlaag bijstellen volgens diezelfde curve. Dit toonde aan dat de relatie niet alleen een correlatie is maar een wezenlijke snelheidscoderingsregel die gedeeld wordt op verschillende niveaus van de bewegingshiërarchie.
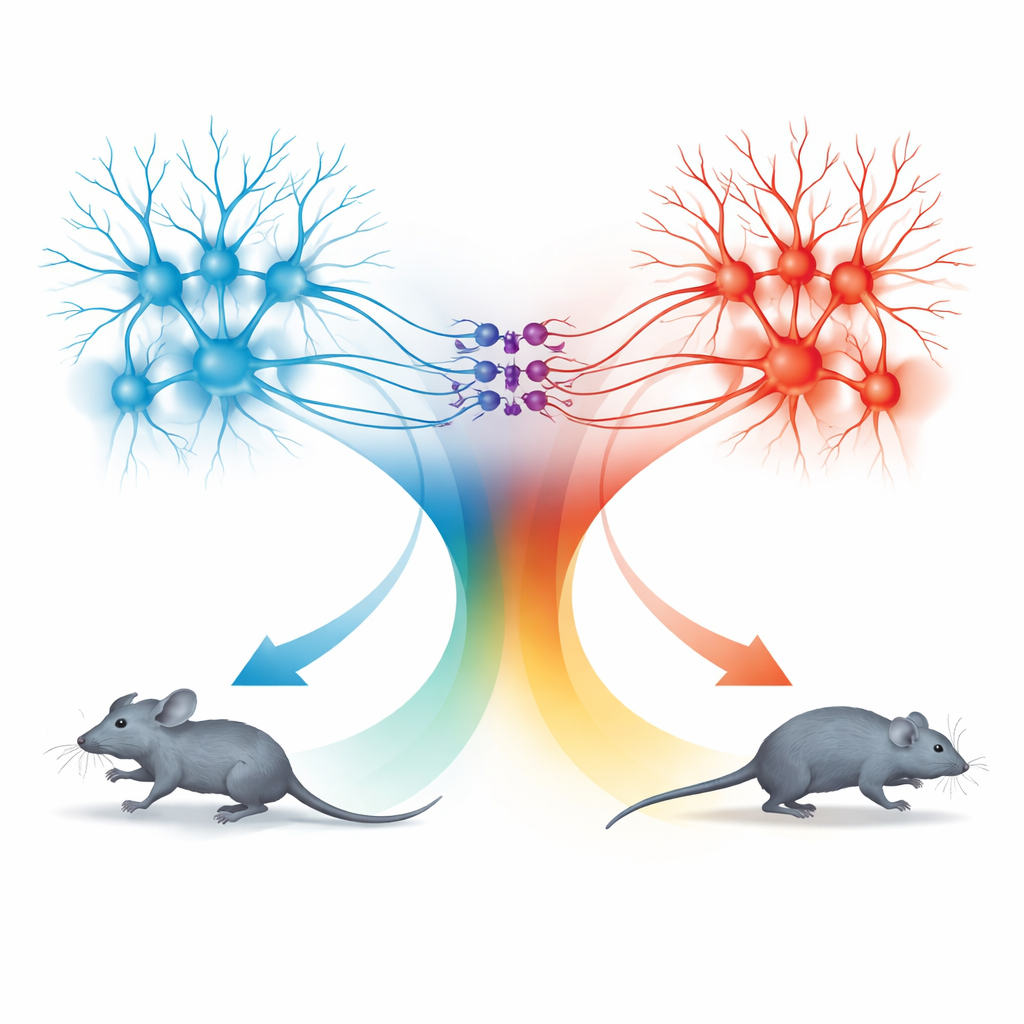
Twee bewegingsunits: vooruit rennen en achteruit terugtrekken
De verrassing kwam uit het onderzoeken van de baan van de superior colliculus naar de dPAG. Neuronen in deze route volgden ook dezelfde snelheidsregel, maar de gedragingen die ze opwekten waren verschillend. Wanneer deze SC-verbonden cellen bij lage lichtfrequenties geactiveerd werden, stopten muizen met bewegen; bij hogere frequenties trokken ze achteruit; en wanneer de stimulatie ophield, braken ze vaak in een korte periode van snel vooruitrennen uit, een zogenaamde rebound-run. Gedetailleerde tracering toonde dat TeA- en SC-signalen grotendeels in gescheiden populaties dPAG-neuronen binnenkwamen. Exciterende cellen gekoppeld aan TeA vormden een "renunit" die rechtlijnig vluchtrennen aandreef, terwijl SC-gekoppelde exciterende cellen een "achteruittrek-unit" vormden. Een kleine set remmende somatostatine (SOM) neuronen in de dPAG verbond deze units in één richting: ze werden geactiveerd door de achteruittrek-unit en onderdrukten op hun beurt de renunit.
Een eenvoudig circuit voor veel verdedigingsstaten
Door selectieve activatie en stillegging te combineren, bracht het team in kaart hoe deze drie celtypen samen vier verschillende gedragingen opleveren: rennen, achteruit trekken, stoppen en rebound-run. Het alleen activeren van TeA-gekoppelde cellen produceerde voorwaarts rennen waarvan de snelheid de gedeelde coderingscurve volgde. Het activeren van SC-gekoppelde cellen zette de achteruittrek-unit en SOM-neuronen aan, schakelde de renunit uit en veroorzaakte afhankelijk van de stimulatiesterkte ofwel stoppen ofwel achteruitbeweging. Wanneer de stimulatie stopte, liet de plotselinge opheffing van SOM-gedreven remming de renunit kort terugveren, wat een uitbarsting van voorwaartse vlucht produceerde. Op deze manier gebruikt de dPAG één kwantitatieve snelheidscode maar leidt die via verschillende microcircuits naar verschillende verdedigingskeuzes.
Wat dit betekent voor het begrijpen van actiekeuzes
Voor niet-specialisten is de kernboodschap dat het brein twee problemen scheidt: hoe krachtig een beweging moet zijn, en welke beweging uitgevoerd moet worden. In dit muismodel zet een gedeelde wiskundige regel neurale activiteit om in bewegingssnelheid, terwijl gespecialiseerde "gedragsunits" in de dPAG beslissen of die snelheid leidt tot vooruit rennen of achteruit terugtrekken. Een kleine remmende schakelbrug maakt het systeem snel wisselen tussen deze toestanden mogelijk, en kan zelfs een rebound-sprint veroorzaken na een periode van opgelegde stilstand. Dit werk levert een concreet voorbeeld van hoe compacte neurale circuits op flexibele wijze complexe overlevingsgedragingen kunnen orkestreren met eenvoudige, herbruikbare coderingsregels.
Bronvermelding: Chen, J., Li, H., Lian, N. et al. A shared speed encoding model for running and backing away behaviours in segregated neural circuits. Nat Commun 17, 4119 (2026). https://doi.org/10.1038/s41467-026-70755-y
Trefwoorden: defensief gedrag, bewegingssnelheid, middenhersencircuits, vluchtreacties, muizenneurowetenschap