Clear Sky Science · fr
Un modèle partagé de codage de la vitesse pour les comportements de fuite et de recul dans des circuits neuronaux séparés
Comment le cerveau fixe le rythme de l’évasion
Quand un animal décide de s’élancer vers l’avant, de se figer ou de reculer prudemment, son cerveau doit non seulement choisir une action mais aussi déterminer la vitesse à laquelle cette action se déroule. Cette étude chez la souris révèle comment une petite région profonde du cerveau calcule la vitesse de déplacement et distribue ce signal de vitesse commun à des réponses défensives très différentes. Le travail offre une fenêtre sur la manière dont le cerveau transforme des sensations vagues de menace en mouvements finement réglés, et explique pourquoi une même menace peut parfois nous pousser à foncer et d’autres fois à reculer ou à nous arrêter.
Un noyau central pour les choix de survie
Au cœur de cette histoire se trouve la substance grise périaqueductale dorsale, ou dPAG, une petite structure du mésencéphale connue depuis longtemps pour son rôle crucial dans les comportements défensifs. Elle reçoit des informations de nombreuses régions cérébrales supérieures et transforme ces signaux en mouvements concrets. Les chercheurs se sont concentrés sur deux sources d’entrée majeures : le cortex d’association temporal (TeA), qui aide à relier les événements sensoriels au contexte, et le colliculus supérieur (SC), une structure qui détecte rapidement les menaces visuelles arrivantes. Les deux régions projettent vers la dPAG, mais il était jusqu’ici peu clair comment leur activité se convertissait en commandes motrices détaillées, comme la vitesse de course.
Des neurones qui prédisent à quelle vitesse vous courrez
En enregistrant finement l’activité de neurones individuels chez des souris éveillées, la tête fixée, courant sur une plateforme tournante, l’équipe a d’abord identifié des cellules dans le TeA dont les taux de décharge augmentaient et diminuaient en fonction de la vitesse de course de l’animal. Ces neurones « liés à la course » ne répondaient pas directement à des flashs visuels, des sons ou des souffles d’air ; au contraire, leur activité montait une à trois secondes avant le début de la course et restait corrélée à la vitesse pendant la course. Lorsque les scientifiques ont tracé le taux de décharge en fonction de la vitesse, ils ont observé que la relation suivait une courbe de saturation : à mesure que la décharge augmentait, la vitesse augmentait rapidement au départ puis se stabilisait vers un maximum, reflétant la limite biologique selon laquelle les animaux ne peuvent pas accélérer indéfiniment.
Un code de vitesse partagé dans une couche cérébrale plus profonde
Le même type d’expérience dans la dPAG a révélé un schéma parallèle. Les neurones y augmentaient aussi leur activité avant et pendant la course, et leur relation activité-vitesse suivait la même courbe mathématique que dans le TeA. Cependant, les neurones de la dPAG étaient plus « efficaces » : pour un même changement d’activité, ils induisaient des variations de vitesse plus importantes, et leur influence se manifestait plus près temporellement du début du mouvement. En stimulant des ensembles spécifiques de neurones TeA et dPAG avec de la lumière à différentes fréquences, les chercheurs ont pu modifier la vitesse de course de manière causale selon cette même courbe. Cela montre que la relation n’est pas seulement une corrélation mais une véritable règle de codage de la vitesse partagée à différents niveaux de la hiérarchie motrice.
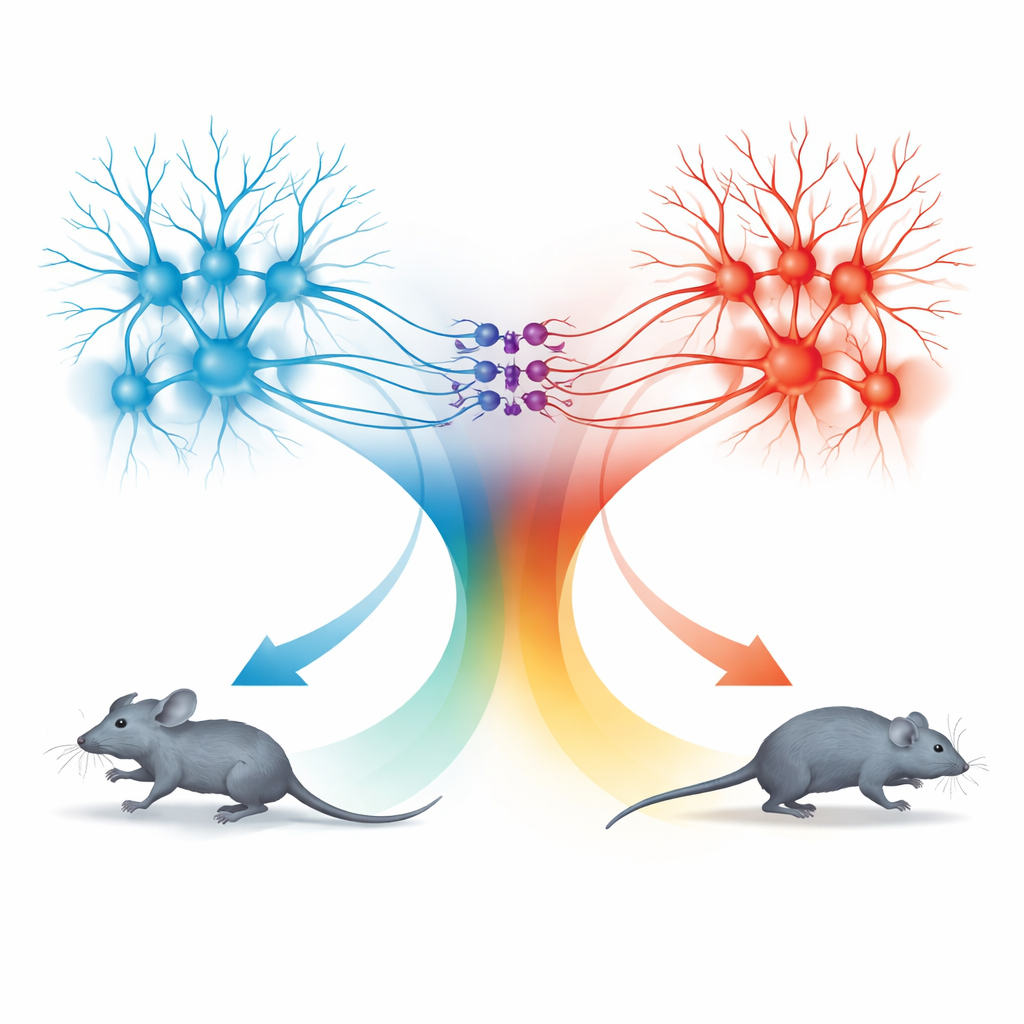
Deux unités de mouvement : courir vers l’avant et reculer
La surprise est venue de l’examen de la voie allant du colliculus supérieur vers la dPAG. Les neurones de cette voie suivaient également la même règle de vitesse, mais les comportements qu’ils induisaient étaient différents. Lorsque ces cellules connectées au SC étaient activées à faibles fréquences lumineuses, les souris s’arrêtaient ; à des fréquences plus élevées, elles reculaient ; et lorsque la stimulation se terminait, elles déclenchaient souvent une brève période de course rapide vers l’avant, appelée course de rebond. Le traçage détaillé a montré que les signaux TeA et SC entraient dans des populations de neurones de la dPAG largement séparées. Les cellules excitatrices liées au TeA formaient une « unité de course » qui provoquait une fuite vers l’avant, tandis que les cellules excitatrices liées au SC formaient une « unité de recul». Un petit ensemble de neurones inhibiteurs exprimant la somatostatine (SOM) dans la dPAG connectait ces unités dans une seule direction : ils étaient activés par l’unité de recul et, à leur tour, suppressaient l’unité de course.
Un circuit simple pour de nombreux états défensifs
En combinant activation et silençage sélectifs, l’équipe a cartographié comment ces trois types cellulaires se combinent pour produire quatre comportements distincts : courir, reculer, s’arrêter et la course de rebond. L’activation uniquement des cellules liées au TeA produisait une course vers l’avant dont la vitesse suivait la courbe de codage partagée. L’activation des cellules liées au SC activait l’unité de recul et les neurones SOM, éteignant l’unité de course et provoquant soit l’arrêt soit le recul selon l’intensité de la stimulation. Lorsque la stimulation cessait, la levée soudaine de l’inhibition portée par les SOM permettait à l’unité de course de rebondir brièvement, produisant un sursaut d’évasion vers l’avant. De cette façon, la dPAG utilise un code quantitatif unique pour la vitesse mais le fait circuler à travers des microcircuits différents afin de générer des choix défensifs distincts.
Ce que cela dit sur le choix d’action
Pour un non-spécialiste, le message clé est que le cerveau sépare deux problèmes : l’intensité d’un mouvement et le choix du mouvement à exécuter. Dans ce modèle de souris, une règle mathématique partagée transforme la décharge neuronale en vitesse de déplacement, tandis que des « unités de comportement » spécialisées dans la dPAG décident si cette vitesse se traduit par une course vers l’avant ou par un recul. Un petit pont inhibiteur permet ensuite au système de basculer rapidement entre ces états, produisant même une ruée de rebond après une période d’immobilité imposée. Ce travail offre un exemple concret de la façon dont des circuits neuronaux compacts peuvent orchestrer de façon flexible des comportements de survie complexes en s’appuyant sur des règles de codage simples et réutilisables.
Citation: Chen, J., Li, H., Lian, N. et al. A shared speed encoding model for running and backing away behaviours in segregated neural circuits. Nat Commun 17, 4119 (2026). https://doi.org/10.1038/s41467-026-70755-y
Mots-clés: comportement défensif, vitesse de déplacement, circuits du mésencéphale, réponses d’évasion, neurosciences chez la souris