Clear Sky Science · pl
Wspólny model kodowania prędkości dla zachowań biegu i cofania się w segregowanych obwodach nerwowych
Jak mózg ustala tempo ucieczki
Kiedy zwierzę decyduje, czy pobiec do przodu, zastygnąć w miejscu czy ostrożnie się cofać, jego mózg musi nie tylko wybrać akcję, lecz także określić, z jaką prędkością ta akcja ma się rozwijać. Badanie na myszach odsłania, jak niewielki obszar głęboko w mózgu oblicza prędkość ruchu i kieruje ten wspólny sygnał prędkości do bardzo różnych reakcji obronnych. Praca daje wgląd w to, jak mózg przekształca nieokreślone poczucie zagrożenia w precyzyjnie dostrojony ruch i dlaczego to samo niebezpieczeństwo czasem popycha nas do natychmiastowego biegu, a innym razem do odwrotu lub zatrzymania.
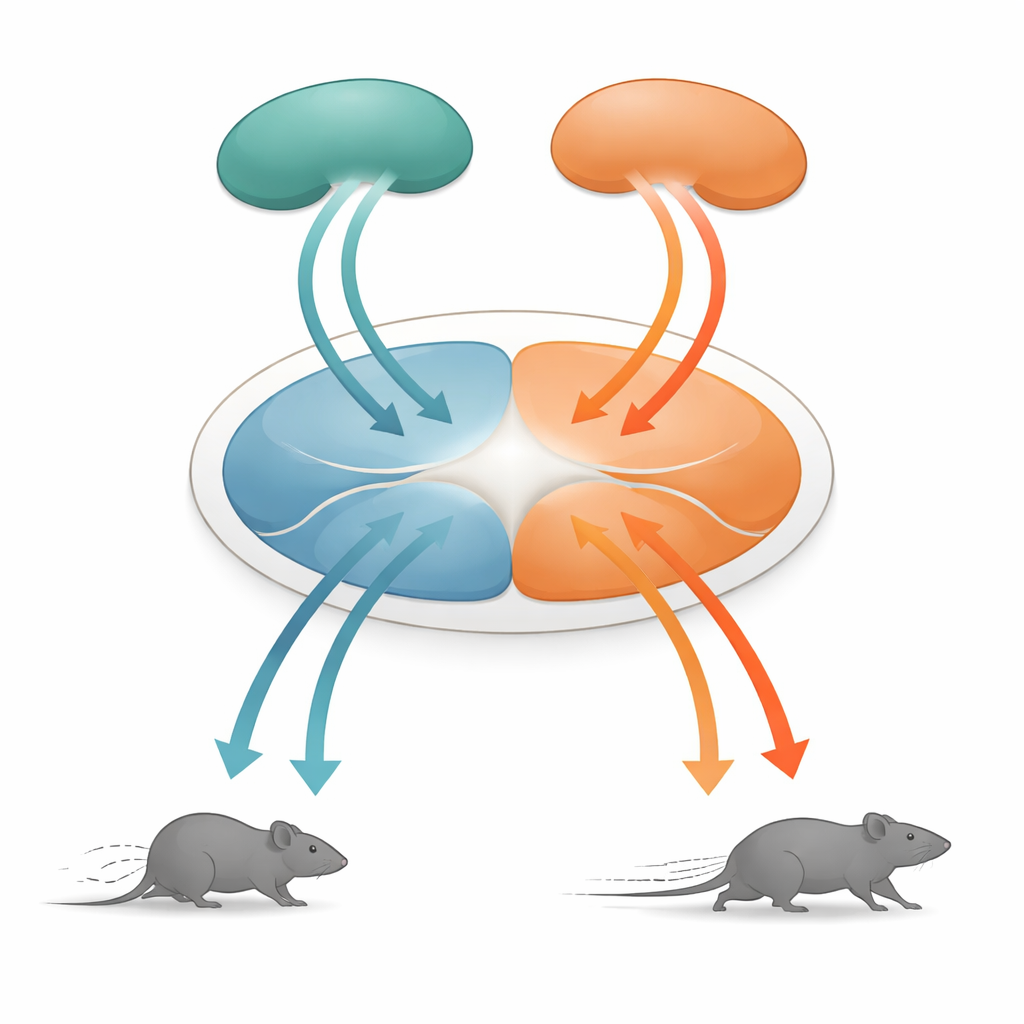
Centralny węzeł wyborów związanych z przetrwaniem
W centrum tej opowieści znajduje się grzbietowa istota szara okołożołożyskowa (dPAG), niewielka struktura w śródmózgowiu od dawna uznawana za kluczową dla zachowań obronnych. Otrzymuje ona informacje z wielu wyższych obszarów mózgu i przekształca te sygnały w konkretne ruchy. Badacze skupili się na dwóch głównych źródłach wejściowych: korze asocjacyjnej skroniowej (TeA), która pomaga łączyć zdarzenia sensoryczne w kontekst, oraz nadkolanku (SC), strukturze szybko wykrywającej zbliżające się zagrożenia wizualne. Oba regiony wysyłają projekcje do dPAG, ale do tej pory nie było jasne, jak ich aktywność przekształca się w szczegółowe polecenia motoryczne, takie jak prędkość biegu.
Neurony, które przewidują, jak szybko pobiegniesz
Wykorzystując delikatne zapisy pojedynczych neuronów u świadomych, unieruchomionych głowowo myszy biegających na talerzu obrotowym, zespół najpierw śledził komórki w TeA, których częstość wyładowań rosła i malała wraz z prędkością biegu zwierzęcia. Te „powiązane z bieganiem” neurony nie reagowały bezpośrednio na błyski, dźwięki czy podmuchy powietrza; zamiast tego ich aktywność narastała na jedną do trzech sekund przed rozpoczęciem biegu i dalej odzwierciedlała prędkość podczas biegu. Gdy naukowcy wykreślili częstość wyładowań względem prędkości biegu, odkryli, że zależność ta podążała za krzywą nasycenia: wraz ze wzrostem aktywności prędkość rośnie szybko na początku, a potem osiąga plateau, odpowiadając biologicznemu ograniczeniu, że zwierzęta nie mogą przyspieszać w nieskończoność.
Wspólny kod prędkości w głębszej warstwie mózgu
Tę samą metodę zastosowano w dPAG i ujawniła podobny wzorzec. Neurony tam także zwiększały aktywność przed i podczas biegu, a zależność między wyładowaniami a prędkością podążała tą samą matematyczną krzywą co w TeA. Jednak neurony dPAG były bardziej „wydajne”: dla danej zmiany w aktywności powodowały większe zmiany prędkości biegu, a ich wpływ ujawniał się bliżej momentu rozpoczęcia ruchu. Pobudzając światłem konkretne zestawy neuronów TeA i dPAG przy różnych częstotliwościach, badacze mogli przyczynowo regulować prędkość biegu zgodnie z tą samą krzywą. To pokazało, że relacja nie jest jedynie korelacją, lecz rzeczywistą regułą kodowania prędkości wspólną dla różnych poziomów hierarchii ruchowej.
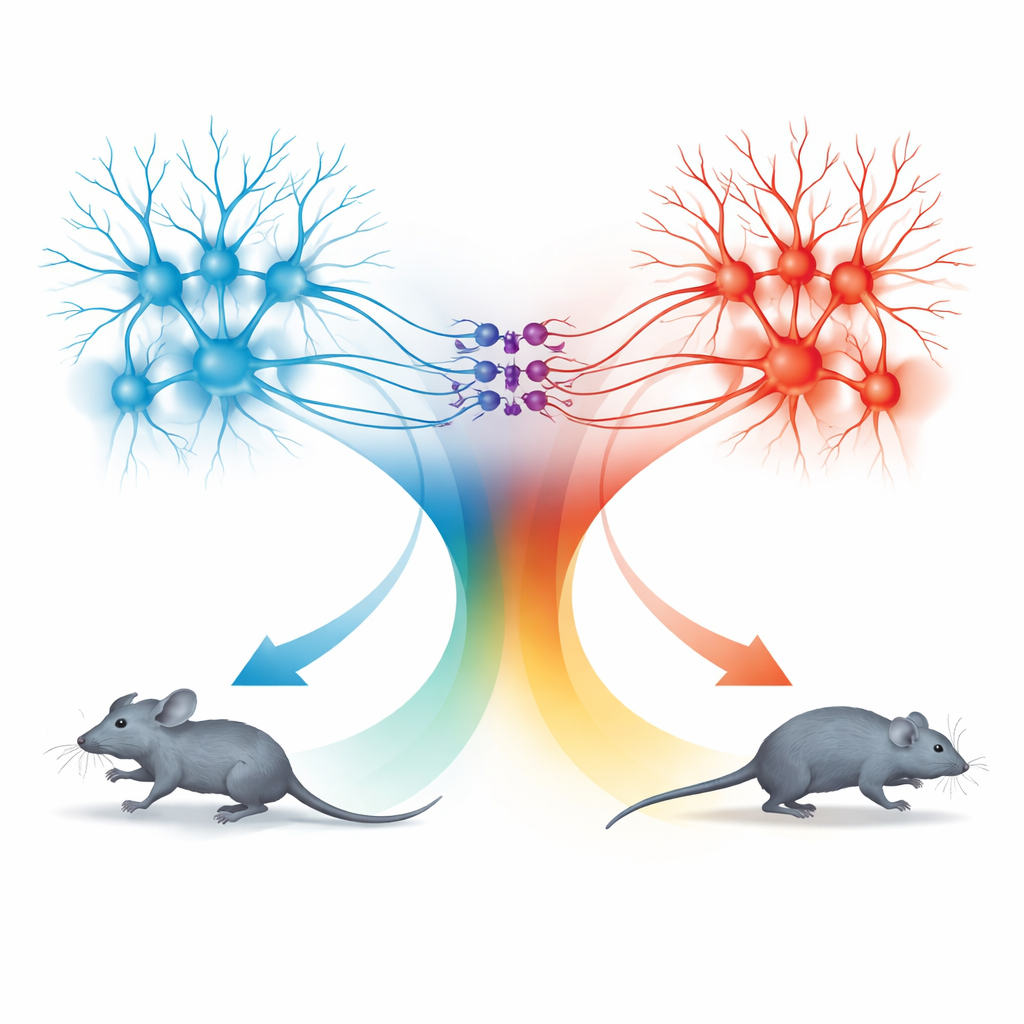
Dwa moduły ruchu: bieganie do przodu i cofanie się
Niespodzianka pojawiła się przy badaniu drogi z nadkolanka do dPAG. Neurony w tej ścieżce również podążały za tą samą regułą prędkości, ale zachowania, które wywoływały, były różne. Gdy komórki połączone z SC były aktywowane przy niskich częstotliwościach światła, myszy przestawały się poruszać; przy wyższych częstotliwościach cofały się; a po zakończeniu stymulacji często wybuchały krótkim okresem szybkiego biegu do przodu, zwanym bieganiem odbiciowym. Szczegółowe mapowanie wykazało, że sygnały z TeA i SC trafiały głównie do oddzielnych populacji neuronów dPAG. Pobudzające komórki powiązane z TeA tworzyły „moduł biegania”, który napędzał prosty ucieczkowy bieg, podczas gdy pobudzające komórki powiązane z SC tworzyły „moduł cofania się”. Niewielka grupa hamujących neuronów somatostatynowych (SOM) w dPAG łączyła te moduły w jednym kierunku: były aktywowane przez moduł cofania i w konsekwencji tłumiły moduł biegania.
Prosty obwód dla wielu stanów obronnych
Łącząc selektywną aktywację i wyciszanie, zespół odwzorował, jak te trzy typy komórek współdziałają, by dać cztery odrębne zachowania: bieganie, cofanie się, zatrzymanie i bieg odbiciowy. Aktywacja samych komórek powiązanych z TeA wywoływała bieg do przodu, którego prędkość podążała za wspólną krzywą kodowania. Aktywacja komórek powiązanych z SC uruchamiała moduł cofania i neurony SOM, wyłączała moduł biegania i powodowała albo zatrzymanie, albo ruch wsteczny w zależności od siły stymulacji. Po ustaniu stymulacji nagłe zniesienie hamowania przez SOM pozwalało modułowi biegania na krótkie odbicie, generując wybuch ucieczki do przodu. W ten sposób dPAG używa jednej ilościowej reguły prędkości, ale kieruje ją przez różne mikroukłady, aby wygenerować odmienne wybory obronne.
Co to znaczy dla zrozumienia wyborów akcji
Dla czytelnika niezaznajomionego ze specjalistycznymi detalami kluczowy wniosek jest taki, że mózg rozdziela dwa problemy: jak silny ma być ruch i jaki ruch wykonać. W tym modelu myszy wspólna reguła matematyczna przekształca aktywność neuronów w prędkość ruchu, podczas gdy wyspecjalizowane „jednostki behawioralne” w dPAG decydują, czy ta prędkość stanie się biegiem do przodu, czy cofaniem się. Mały hamujący mostek pozwala systemowi szybko przełączać się między tymi stanami, a nawet wywołać odbiciowy pęd po okresie wymuszonego bezruchu. Praca ta daje konkretne przykłady, jak zwarte obwody nerwowe mogą elastycznie orkiestrująć złożone zachowania przetrwania, stosując proste, wielokrotnego użytku reguły kodowania.
Cytowanie: Chen, J., Li, H., Lian, N. et al. A shared speed encoding model for running and backing away behaviours in segregated neural circuits. Nat Commun 17, 4119 (2026). https://doi.org/10.1038/s41467-026-70755-y
Słowa kluczowe: zachowanie obronne, prędkość ruchu, obwody śródmózgowia, reakcje ucieczki, neuronauka myszy