Clear Sky Science · it
Un modello condiviso di codifica della velocità per i comportamenti di corsa e di indietreggiamento in circuiti neuronali segregati
Come il cervello fissa il ritmo della fuga
Quando un animale decide se scattare in avanti, restare immobile o indietreggiare con cautela, il suo cervello non deve solo scegliere un’azione ma anche determinare la velocità con cui quell’azione si svolgerà. Questo studio nei topi svela come una piccola regione profonda del cervello calcoli la velocità del movimento e indirizzi quel segnale di velocità condiviso verso risposte difensive molto diverse. Il lavoro offre una finestra su come il cervello trasformi sensazioni vaghe di minaccia in movimenti precisamente calibrati, e perché lo stesso pericolo a volte ci spinga ad avanzare di corsa e altre volte a ritirarci o fermarci.
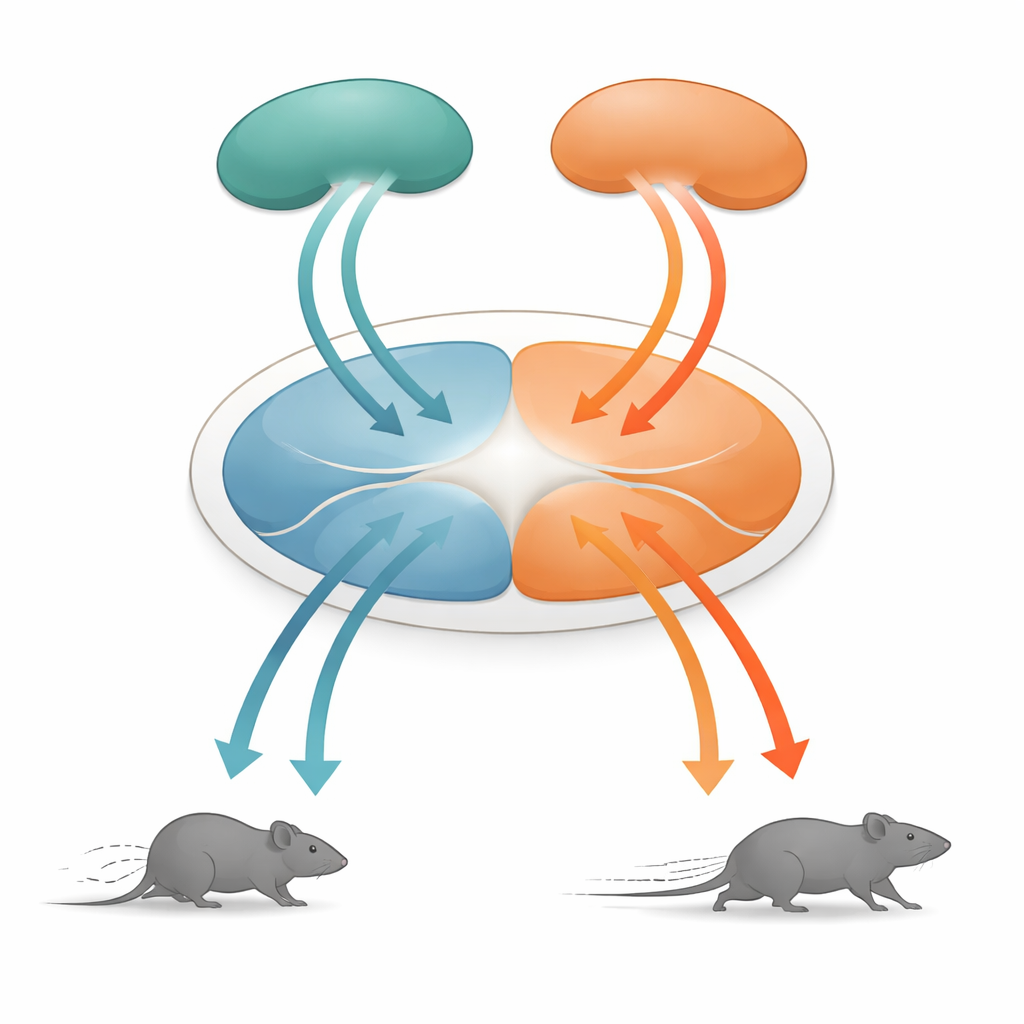
Un hub centrale per le scelte di sopravvivenza
Al centro di questa storia c’è la grigia periacqueduttale dorsale, o dPAG, una piccola struttura del mesencefalo da tempo nota per essere cruciale nei comportamenti difensivi. Riceve informazioni da molte aree corticali superiori e trasforma quei segnali in movimenti concreti. I ricercatori si sono concentrati su due principali sorgenti di input: la corteccia associativa temporale (TeA), che aiuta a collegare eventi sensoriali al contesto, e il collicolo superiore (SC), una struttura che rileva rapidamente minacce visive in avvicinamento. Entrambe le regioni inviano proiezioni alla dPAG, ma finora non era chiaro come la loro attività venisse trasformata in comandi motori dettagliati come la velocità di corsa.
Neuroni che predicono quanto velocemente correrai
Usando registrazioni delicate da singoli neuroni in topi svegli, con la testa immobilizzata e che correvano su una piattaforma rotante, il gruppo ha prima tracciato cellule nella TeA i cui tassi di scarica aumentavano e diminuivano con la velocità di corsa dell’animale. Questi neuroni «relativi alla corsa» non rispondevano direttamente a lampi, suoni o getti d’aria; invece la loro attività aumentava da uno a tre secondi prima che l’animale iniziasse a correre e continuava a rispecchiare la sua velocità durante la corsa. Quando gli scienziati tracciarono il tasso di scarica in funzione della velocità, osservarono che la relazione seguiva una curva di saturazione: all’aumentare della scarica, la velocità saliva rapidamente all’inizio per poi stabilizzarsi a un massimo, coerente con il limite biologico che gli animali non possono accelerare indefinitamente.
Un codice di velocità condiviso in uno strato cerebrale più profondo
Lo stesso tipo di esperimento nella dPAG rivelò uno schema parallelo. Anche i neuroni lì aumentavano la loro attività prima e durante la corsa, e la loro relazione scarica-velocità seguiva la stessa curva matematica osservata nella TeA. Tuttavia, i neuroni della dPAG erano più «efficienti»: per una data variazione di scarica producevano variazioni maggiori nella velocità di corsa, e la loro influenza si attivava più vicino temporalmente all’inizio del movimento. Stimolando con luce a diverse frequenze insiemi specifici di neuroni della TeA e della dPAG, i ricercatori poterono modulare causalmente la velocità di corsa in accordo con quella stessa curva. Ciò dimostrò che la relazione non è solo una correlazione ma una vera regola di codifica della velocità condivisa tra livelli della gerarchia motoria.
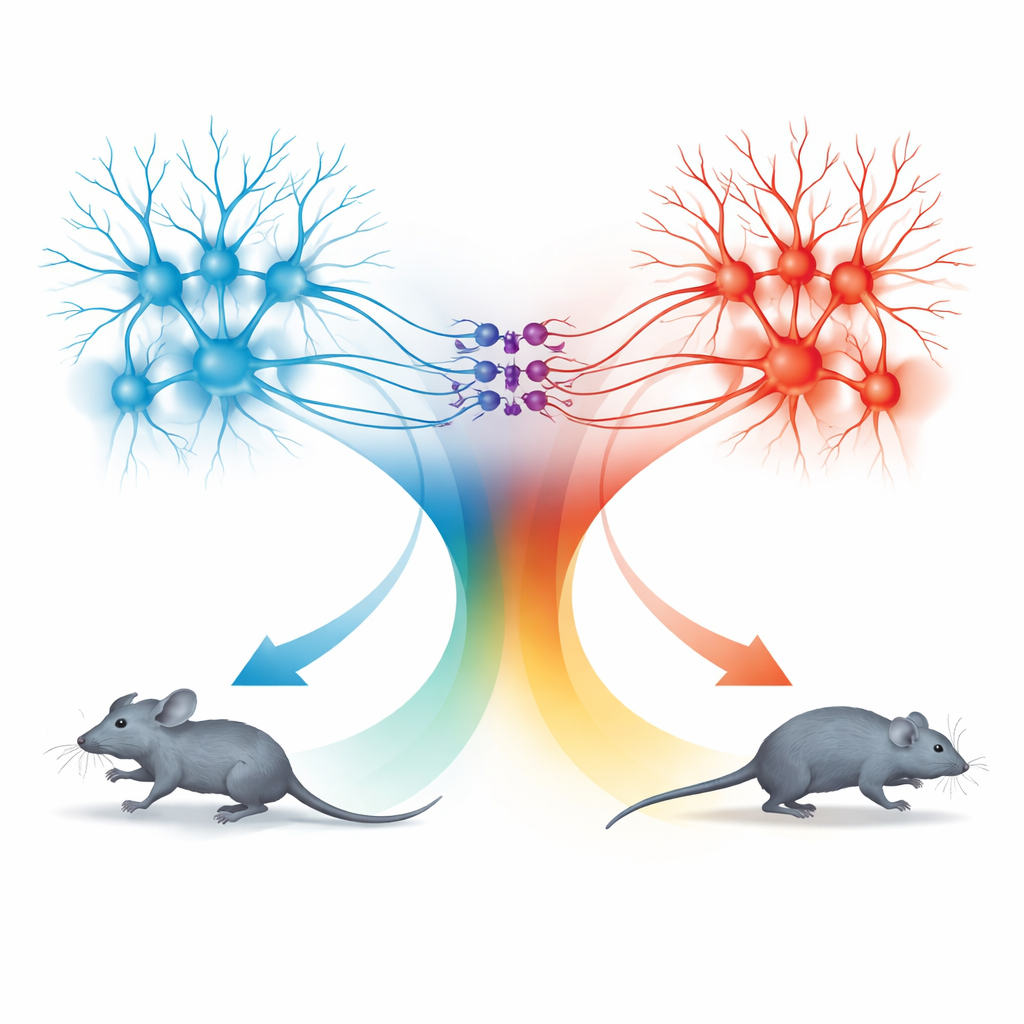
Due unità di movimento: corsa in avanti e indietreggiamento
La sorpresa emerse dall’esame della via che dal collicolo superiore arriva alla dPAG. I neuroni in questo percorso seguivano anch’essi la stessa regola di velocità, ma i comportamenti che evocavano erano diversi. Quando queste cellule connesse al SC venivano attivate a basse frequenze luminose, i topi smettevano di muoversi; a frequenze più alte indietreggiavano; e quando la stimolazione terminava spesso esplodevano in un breve periodo di corsa rapida in avanti, chiamato corsa di rimbalzo. L’analisi dettagliata delle connessioni mostrò che i segnali di TeA e SC entravano in popolazioni per lo più separate di neuroni nella dPAG. Le cellule eccitatorie collegate alla TeA formavano un’«unità di corsa» che guidava la fuga in avanti diretta, mentre le cellule eccitatorie collegate al SC costituivano un’«unità di indietreggiamento». Un piccolo gruppo di neuroni inibitori che esprimono somatostatina (SOM) nella dPAG collegava questi due blocchi in una sola direzione: venivano attivati dall’unità di indietreggiamento e a loro volta sopprimevano l’unità di corsa.
Un circuito semplice per molti stati difensivi
Combinando attivazione e silenziamento selettivi, il team mappò come questi tre tipi cellulari si combinano per produrre quattro comportamenti distinti: corsa, indietreggiare, fermarsi e corsa di rimbalzo. L’attivazione delle sole cellule collegate alla TeA generava corsa in avanti la cui velocità seguiva la curva di codifica condivisa. L’attivazione delle cellule collegate al SC attivava l’unità di indietreggiamento e i neuroni SOM, spegnendo l’unità di corsa e provocando o l’arresto o il moto all’indietro a seconda dell’intensità della stimolazione. Quando la stimolazione cessava, il rapido sollevamento dell’inibizione guidata da SOM permetteva all’unità di corsa di rimbalzare brevemente, producendo un’esplosione di fuga in avanti. In questo modo la dPAG utilizza un unico codice quantitativo della velocità ma lo instrada attraverso microcircuiti diversi per generare scelte difensive differenti.
Cosa significa per la comprensione delle scelte d’azione
Per un lettore non specialista, il messaggio chiave è che il cervello separa due problemi: quanto vigoroso deve essere un movimento e quale movimento eseguire. In questo modello murino, una regola matematica condivisa trasforma la scarica neuronale in velocità del movimento, mentre «unità comportamentali» specializzate nella dPAG decidono se quella velocità si tradurrà in corsa in avanti o in indietreggiamento. Un piccolo ponte inibitorio consente poi al sistema di passare rapidamente tra questi stati, arrivando persino a produrre una scatto di rimbalzo dopo un periodo di immobilità forzata. Questo lavoro offre un esempio concreto di come circuiti neurali compatti possano orchestrare in modo flessibile comportamenti di sopravvivenza complessi utilizzando regole di codifica semplici e riutilizzabili.
Citazione: Chen, J., Li, H., Lian, N. et al. A shared speed encoding model for running and backing away behaviours in segregated neural circuits. Nat Commun 17, 4119 (2026). https://doi.org/10.1038/s41467-026-70755-y
Parole chiave: comportamento difensivo, velocità del movimento, circuiti del mesencefalo, risposte di fuga, neuroscienze del topo