Clear Sky Science · de
Ein gemeinsames Geschwindigkeitskodierungsmodell für Vorwärtslaufen und Rückwärtsweichen in segregierten neuronalen Schaltkreisen
Wie das Gehirn das Tempo für die Flucht vorgibt
Wenn ein Tier entscheidet, ob es vorpreschen, erstarren oder vorsichtig zurückweichen soll, muss sein Gehirn nicht nur eine Handlung auswählen, sondern auch festlegen, wie schnell diese Handlung abläuft. Diese Studie an Mäusen zeigt, wie eine winzige Region tief im Gehirn die Bewegungsgeschwindigkeit berechnet und dieses gemeinsame Geschwindigkeitssignal in sehr unterschiedliche Abwehrreaktionen einspeist. Die Arbeit bietet Einblick darin, wie das Gehirn vage Bedrohungsgefühle in präzise abgestimmte Bewegungen übersetzt und warum dieselbe Gefahr uns manchmal vorwärts stürzen und ein anderes Mal zurückweichen oder stoppen lässt.
Ein zentrales Schaltzentrum für Überlebensentscheidungen
Im Mittelpunkt dieser Geschichte steht die dorsale Periaquäduktalgrau-Region, kurz dPAG, eine kleine Struktur im Mittelhirn, die seit langem als entscheidend für Abwehrverhalten bekannt ist. Sie erhält Informationen aus vielen höheren Hirnbereichen und verwandelt diese Signale in konkrete Bewegungen. Die Forscher konzentrierten sich auf zwei wichtige Eingangsquellen: den temporalen Assoziationskortex (TeA), der dabei hilft, Sinnesereignisse in ihren Kontext einzubetten, und die superiore Colliculus (SC), eine Struktur, die herannahende visuelle Bedrohungen schnell erkennt. Beide Regionen senden Projektionen in die dPAG, doch war bislang unklar, wie ihre Aktivität in detaillierte motorische Befehle wie Laufgeschwindigkeit umgesetzt wird.
Neuronen, die vorhersagen, wie schnell das Tier laufen wird
Mithilfe empfindlicher Aufzeichnungen einzelner Neuronen in wachen, am Kopf fixierten Mäusen, die sich auf einer Drehscheibe bewegten, verfolgte das Team zunächst Zellen im TeA, deren Feuerraten mit der Laufgeschwindigkeit des Tieres anstiegen und fielen. Diese „laufbezogenen" Neuronen reagierten nicht direkt auf Lichtblitze, Geräusche oder Luftstöße; stattdessen stieg ihre Aktivität ein bis drei Sekunden bevor das Tier zu laufen begann an und spiegelte während des Laufs seine Geschwindigkeit wider. Als die Wissenschaftler die Feuerrate gegen die Laufgeschwindigkeit auftrugen, zeigte sich eine sättigende Kurve: Mit zunehmender Feuerrate stieg die Geschwindigkeit zunächst schnell an und glättete sich dann bei einem Maximum, was dem biologischen Limit entspricht, dass Tiere nicht unendlich beschleunigen können.
Ein gemeinsamer Geschwindigkeitscode in einer tieferen Hirnschicht
Dasselbe Experiment im dPAG offenbarte ein paralleles Muster. Auch dort erhöhten Neuronen ihre Feuerrate vor und während des Laufens, und ihre Feuerrate-zu-Geschwindigkeit-Beziehung folgte derselben mathematischen Kurve wie im TeA. Allerdings waren dPAG-Neuronen „effizienter": Bei derselben Änderung der Feuerrate bewirkten sie größere Änderungen der Laufgeschwindigkeit, und ihr Einfluss setzte näher am Bewegungsbeginn ein. Durch gezielte Stimulation spezifischer Gruppen von TeA- und dPAG-Neuronen mit Licht in unterschiedlichen Frequenzen konnten die Forscher die Laufgeschwindigkeit kausal entlang dieser Kurve hoch- und herunterregeln. Das zeigte, dass die Beziehung nicht nur eine Korrelation ist, sondern eine echte, über Ebenen der Bewegungshierarchie geteilte Geschwindigkeitscodierungsregel.
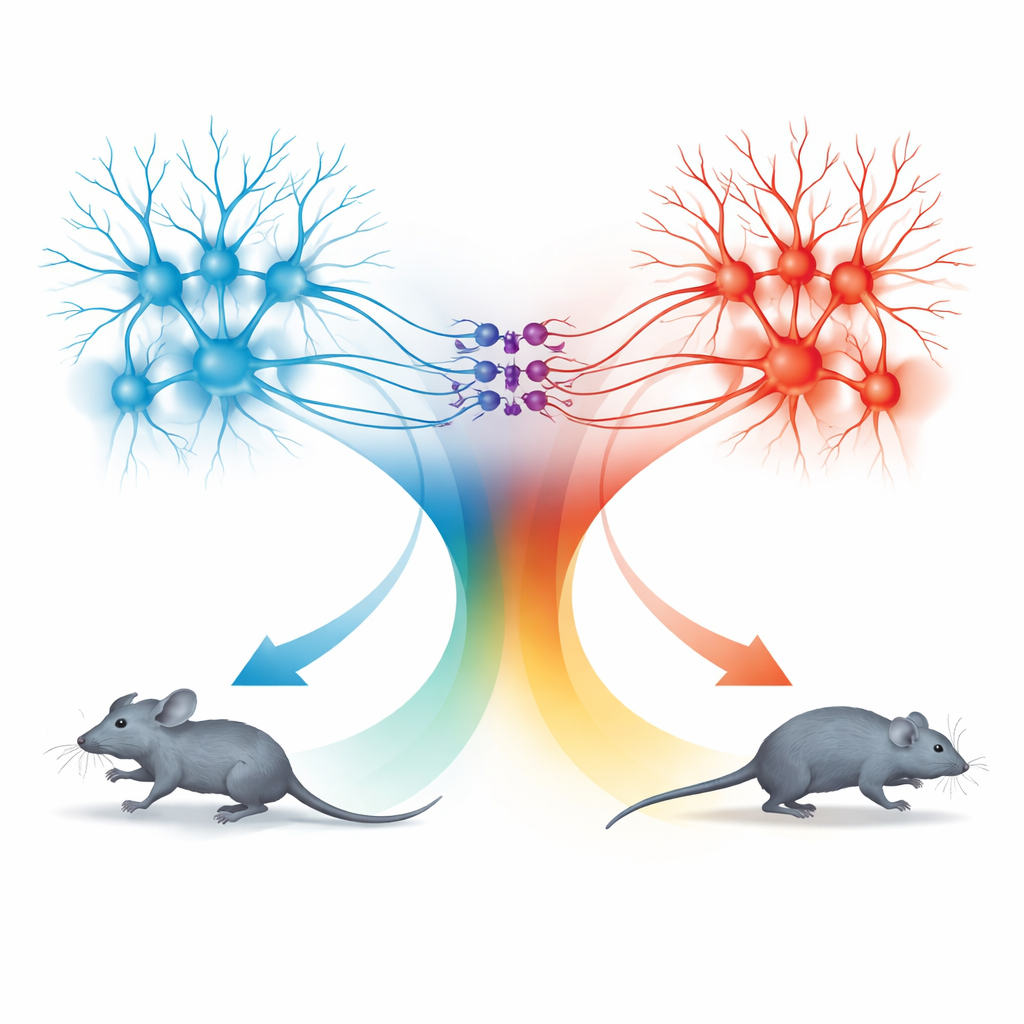
Zwei Bewegungseinheiten: Vorwärtslaufen und Zurückweichen
Die Überraschung kam bei der Untersuchung des Pfades vom superioren Colliculus in die dPAG. Neuronen auf diesem Weg folgten ebenfalls derselben Geschwindigkeitsregel, doch die von ihnen ausgelösten Verhaltensweisen waren unterschiedlich. Wenn diese SC-verbundenen Zellen mit niedrigen Lichtfrequenzen aktiviert wurden, hörten die Mäuse auf sich zu bewegen; bei höheren Frequenzen wichen sie zurück; und wenn die Stimulation endete, brachen sie oft in eine kurze Phase schnellen Vorwärtslaufens aus, die als Rebound-Lauf bezeichnet wird. Detailliertes Tracing zeigte, dass TeA- und SC-Signale weitgehend getrennte Populationen von dPAG-Neuronen erreichten. TeA-verbundene exzitatorische Zellen bildeten eine „Laufeinheit", die geradliniges Fluchtlaufen antrieb, während SC-verbundene exzitatorische Zellen eine „Zurückweich-Einheit" bildeten. Eine kleine Gruppe inhibitorischer Somatostatin-(SOM-)Neuronen in der dPAG verband diese Einheiten in eine Richtung: Sie wurden von der Zurückweich-Einheit aktiviert und unterdrückten im Gegenzug die Laufeinheit.
Ein einfacher Schaltkreis für viele Abwehrzustände
Durch Kombination selektiver Aktivierung und Stilllegung kartierte das Team, wie diese drei Zelltypen zusammenwirken, um vier unterschiedliche Verhaltensweisen zu erzeugen: Laufen, Zurückweichen, Stoppen und Rebound-Laufen. Die alleinige Aktivierung der TeA-verbundenen Zellen erzeugte Vorwärtslauf, dessen Geschwindigkeit der gemeinsamen Kodierungskurve folgte. Die Aktivierung der SC-verbundenen Zellen schaltete die Zurückweich-Einheit und die SOM-Neuronen ein, schaltete die Laufeinheit ab und führte je nach Stimulationsstärke zu Stoppen oder Rückwärtsbewegung. Wenn die Stimulation endete, erlaubte das plötzliche Aufheben der SOM-vermittelten Hemmung der Laufeinheit einen kurzen Rebound, der einen Ausbruch von Vorwärtsflucht hervorrief. Auf diese Weise nutzt die dPAG einen einzigen quantitativen Geschwindigkeitscode, leitet ihn aber durch unterschiedliche Mikroschaltkreise, um verschiedene Abwehrentscheidungen zu erzeugen.
Was das für das Verständnis von Handlungsentscheidungen bedeutet
Für Nicht-Spezialisten ist die Kernbotschaft, dass das Gehirn zwei Probleme trennt: wie stark eine Bewegung sein sollte und welche Bewegung ausgeführt werden soll. In diesem Mausmodell verwandelt eine gemeinsame mathematische Regel neuronales Feuern in Bewegungsgeschwindigkeit, während spezialisierte „Verhaltenseinheiten" in der dPAG entscheiden, ob diese Geschwindigkeit in Vorwärtslaufen oder Zurückweichen umgesetzt wird. Eine kleine inhibitorische Brücke erlaubt es dem System dann, schnell zwischen diesen Zuständen zu wechseln und sogar nach einer erzwungenen Stillstandsphase einen Ausbruch zu produzieren. Diese Arbeit liefert ein konkretes Beispiel dafür, wie kompakte neuronale Schaltkreise komplexe Überlebensverhalten flexibel mit einfachen, wiederverwendbaren Kodierungsregeln orchestrieren können.
Zitation: Chen, J., Li, H., Lian, N. et al. A shared speed encoding model for running and backing away behaviours in segregated neural circuits. Nat Commun 17, 4119 (2026). https://doi.org/10.1038/s41467-026-70755-y
Schlüsselwörter: Abwehrverhalten, Bewegungsgeschwindigkeit, Mittelhirnschaltkreise, Fluchtreaktionen, Maus-Neurowissenschaft