Clear Sky Science · ar
نموذج ترميز سرعة مشترك لسلوكيات الجري والتراجع في دوائر عصبية مفصولة
كيف يضبط الدماغ وتيرة الهروب
عندما يقرر الحيوان ما إذا كان سيندفع للأمام بسرعة، أو يتجمد، أو يتراجع بحذر، يجب على دماغه ألا يختار الفعل فحسب، بل أيضاً أن يحدد السرعة التي يتكشف بها ذلك الفعل. تكتشف هذه الدراسة في الفئران كيف أن منطقة صغيرة عميقة في الدماغ تحسب سرعة الحركة وتوجه إشارة السرعة المشتركة تلك نحو استجابات دفاعية مختلفة تماماً. العمل يوفّر نافذة على كيفية تحويل الدماغ لمشاعر التهديد المبهمة إلى حركة مضبوطة بدقة، ولماذا قد تدفعنا نفس الخطورة أحياناً إلى الاندفاع للأمام وأحياناً أخرى إلى التراجع أو التوقف.
مركز محوري لخيارات البقاء
في قلب هذه القصة تقع المنطقة الرمادية المحيطة بالقناة الظهرية، أو dPAG، وهي بنية صغيرة في الدماغ الأوسط معروفة منذ زمن بعيد بأهميتها في السلوكيات الدفاعية. تستقبل معلومات من العديد من المناطق العليا في الدماغ وتحوّل تلك الإشارات إلى حركات ملموسة. ركز الباحثون على مصدرين رئيسيين للمدخلات: القشرة الترابطية الصدغية (TeA)، التي تساعد على ربط الأحداث الحسية بالسياق، والمَخَرّ الأعلى (SC)، وهي بنية تكتشف بسرعة التهديدات البصرية الوشيكة. كلا المنطقتين ترسلان عُصبونات إلى dPAG، لكن حتى الآن لم تكن واضحة كيفية تحويل نشاطهما إلى أوامر حركية مفصّلة مثل سرعة الجري.
عصبونات تتنبأ بمدى سرعتك عند الجري
باستخدام تسجيلات حساسة من عصبونات فردية في فئران يقظة وملتصقة الرأس تجري على طاولة دوّارة، تتبّع الفريق أولاً خلايا في TeA ارتفعت وانخفضت معدلات إطلاقها مع سرعة جري الحيوان. هذه الخلايا «المتعلقة بالجري» لم تتجاوب مباشرة مع ومضات ضوء أو أصوات أو نفخات هواء؛ بل ارتفعت إطلاقاتها قبل ثانية إلى ثلاث ثوانٍ من بدء الجري واستمرت في مراعاة سرعته أثناء الجري. عندما رسم العلماء منحنى معدل الإطلاق مقابل سرعة الجري، وجدوا أن العلاقة اتبعت منحنى تشبّع: مع ازدياد الإطلاق زادت السرعة سريعاً في البداية ثم استقرت عند قيمة قصوى، متطابقة مع الحد البيولوجي الذي يمنع الحيوانات من التسارع إلى ما لا نهاية.
رمز سرعة مشترك في طبقة أعمق من الدماغ
أظهر نفس نوع التجربة في dPAG نمطاً موازياً. فالعصبونات هناك أيضاً زادت إطلاقها قبل وأثناء الجري، وكانت علاقة الإطلاق بالسرعة تتبع نفس المعادلة الرياضية كما في TeA. ومع ذلك، كانت عصبونات dPAG أكثر «كفاءة»: لتغيير معين في الإطلاق، أحدثت تغييرات أكبر في سرعة الجري، وتأثيرها بدأ أقرب زمنياً لبدء الحركة. من خلال تحفيز مجموعات محددة من عصبونات TeA وdPAG بالضوء بترددات مختلفة، استطاع الباحثون ضبط سرعة الجري تصاعدياً وتنازلياً بما يتوافق مع ذلك المنحنى نفسه. هذا أظهر أن العلاقة ليست مجرد ارتباط بل قاعدة ترميز سرعة حقيقية مشتركة عبر مستويات هرمية الحركة.
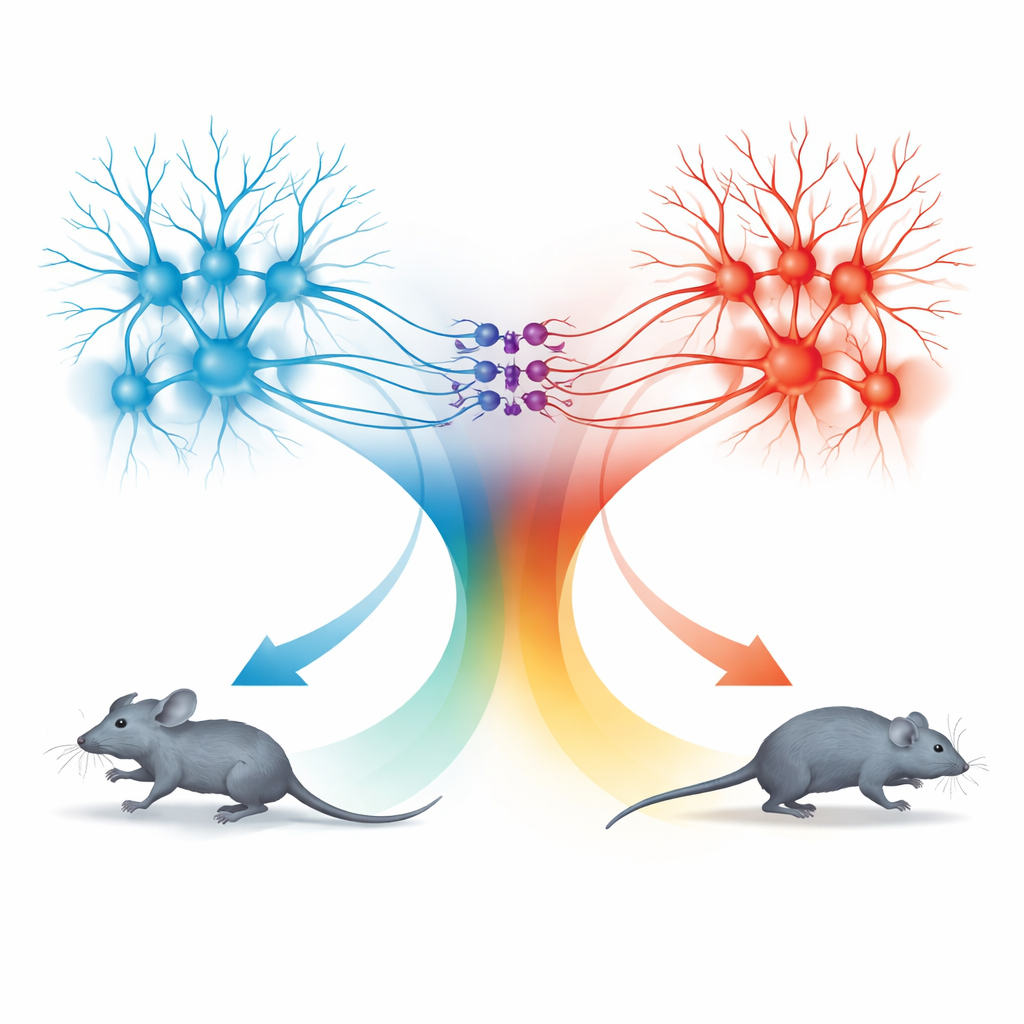
وحدتا حركة: الجري للأمام والتراجع للخلف
المفاجأة جاءت من فحص المسار من المَخَرّ الأعلى إلى dPAG. العصبونات في هذا المسار اتبعت أيضاً قاعدة السرعة نفسها، لكن السلوكيات التي حرضتها كانت مختلفة. عندما نُشِّطت هذه الخلايا المتصلة بـSC بترددات ضوئية منخفضة، توقفت الفئران عن الحركة؛ عند ترددات أعلى تراجعت إلى الخلف؛ وعندما انتهى التحفيز، غالباً ما اندفعت فجأة في فترة قصيرة من الجري الأمامي السريع، سميت بالجري الارتدادي. أظهر التتبع التفصيلي أن إشارات TeA وSC دخلت مجموعات منفصلة إلى حد كبير من عصبونات dPAG. شكلت الخلايا المثيرة للارتباط بـTeA «وحدة جري» تقود هروباً أمامياً مباشراً، بينما شكلت الخلايا المثيرة المرتبطة بـSC «وحدة تراجع». مجموعة صغيرة من الخلايا المثبطة المنتجة للسوماتوستاتين (SOM) في dPAG ربطت هذه الوحدات في اتجاه واحد: كانت تُفَعَّل بواسطة وحدة التراجع وتقوم بدورها بكبح وحدة الجري.
دائرة بسيطة لعدة حالات دفاعية
بدمج التفعيل والكمون الانتقائي، رسم الفريق خريطة كيفية تفاعل هذه الأنواع الثلاثة من الخلايا لإنتاج أربع سلوكيات مميزة: الجري، والتراجع، والتوقف، والجري الارتدادي. تفعيل خلايا مرتبطة بـTeA بمفردها أنتج جريًا أمامياً تتبع سرعته منحنى الترميز المشترك. تفعيل خلايا مرتبطة بـSC أدار وحدة التراجع وخلايا SOM، مما أوقف وحدة الجري وتسبب إما بالتوقف أو بالحركة للخلف اعتماداً على قوة التحفيز. عند توقف التحفيز، أدت الزيادة المفاجئة في رفع كبح SOM إلى السماح لوحدة الجري بالارتداد مؤقتاً، مولدة اندفاعة هروب أمامية. بهذه الطريقة، يستخدم dPAG قاعدة كمية واحدة للسرعة لكنه يمررها عبر ميكروسيركيتات مختلفة لتوليد خيارات دفاعية مختلفة.
ماذا يعني هذا لفهم اختيار الحركة
لغير المتخصصين، الرسالة الأساسية هي أن الدماغ يفصل بين مشكلتين: مدى قوة الحركة التي ينبغي أن تكون، وأي حركة ينبغي تنفيذها. في نموذج الفأر هذا، تتحول قاعدة رياضية مشتركة إطلاقات العصبونات إلى سرعة حركة، بينما تقرر «وحدات السلوك» المتخصصة في dPAG ما إذا كانت تلك السرعة ستتحول إلى جري للأمام أو تراجع للخلف. جسر مثبط صغير يسمح للنظام بالتبديل بسرعة بين هذه الحالات، بل وإنتاج انطلاق ارتدادي بعد فترة من الثبات المفروض. يقدم هذا العمل مثالاً ملموساً على كيف يمكن لدارات عصبية مدمجة أن تنسق بمرونة سلوكيات بقاء معقدة باستخدام قواعد ترميز بسيطة وقابلة لإعادة الاستخدام.
الاستشهاد: Chen, J., Li, H., Lian, N. et al. A shared speed encoding model for running and backing away behaviours in segregated neural circuits. Nat Commun 17, 4119 (2026). https://doi.org/10.1038/s41467-026-70755-y
الكلمات المفتاحية: سلوك دفاعي, سرعة الحركة, دارات الدماغ الأوسط, استجابات الهروب, علم أعصاب الفأر