Clear Sky Science · zh
硫氧还蛋白及其伴侣蛋白对密苏里放线菌游动孢子鞭毛形成至关重要
某些细菌如何按需构建微小尾巴
许多细菌通过长而鞭状的尾巴——鞭毛来移动。对一种土壤微生物Actinoplanes missouriensis而言,恰在其所需时刻构建这些尾巴关系重大:当雨水或水体出现时,其孢子必须瞬间变为游动体。本文揭示了一对小型蛋白如何像分子开关一样控制何时建造这些游动尾巴。
从静默孢子到突然游动
A. missouriensis通常以土壤中的分枝菌丝体形式生长。在干旱条件下,它会在菌丝末端形成称为孢囊(sporangia)的圆形结构。在每个孢囊内,数百个圆状孢子形成并为外界生活做准备。当水到来时,孢囊打开,将孢子释放到液体中。这些孢子迅速长出鞭毛,成为可短暂游动的“游动孢子”,帮助它们在定居生长前扩散到新地点。由于这一游动阶段短暂且时机严格受控,细胞必须精确调节鞭毛组装的时机和方式。
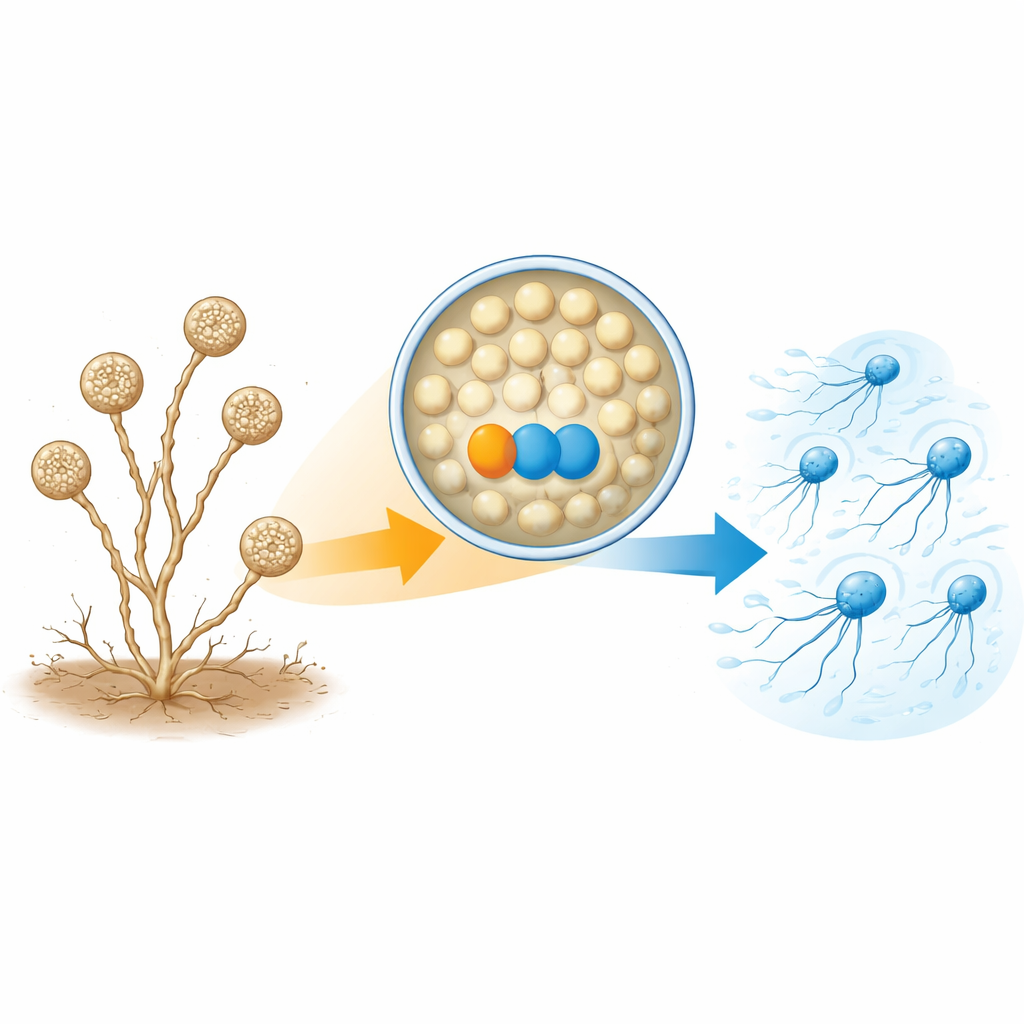
构建尾巴所需的一对蛋白
作者将注意力集中在两种在游动孢子中丰度远高于萌发细胞的蛋白上。一种是称为TrxA的硫氧还蛋白,另一种是其伴侣蛋白PtxA。研究者删除任一TrxA或PtxA基因后,孢子在孢囊内仍能正常形成并按时释放——但它们无法游动。电子显微镜显示了原因:这些突变体的大多数孢子根本缺乏鞭毛,或仅有少数短鞭毛。然而,鞭毛基因的信使RNA水平以及至少一种关键鞭毛构件蛋白(FliC)的量基本正常。这表明TrxA和PtxA并不负责开启或关闭鞭毛基因;相反,它们对鞭毛结构的实际组装是必需的。
一种经典氧化还原蛋白的非氧化还原工作
硫氧还蛋白通常通过其一对半胱氨酸残基来重排其他蛋白中的二硫键以发挥作用。为测试这种典型的氧化还原活性是否在此处必需,团队纯化了TrxA,并在试管实验中表明其表现如同典型硫氧还蛋白:能还原标准蛋白底物。当他们改变一或两个关键半胱氨酸时,TrxA失去了这种氧化还原能力。令人惊讶的是,携带这些“氧化还原失活”版本TrxA的细菌仍能产生完全带鞭毛、可运动的孢子。相反,用另一种细菌中最相似的硫氧还蛋白替换TrxA的部分序列并不能恢复鞭毛形成,即便替代蛋白具有正常的硫氧还蛋白活性。通过系统地交换两者之间的区域,作者将关键特征缩小到TrxA中的一段短小的五个氨基酸序列EKVEQ,该序列在许多Actinoplanes物种中保守。
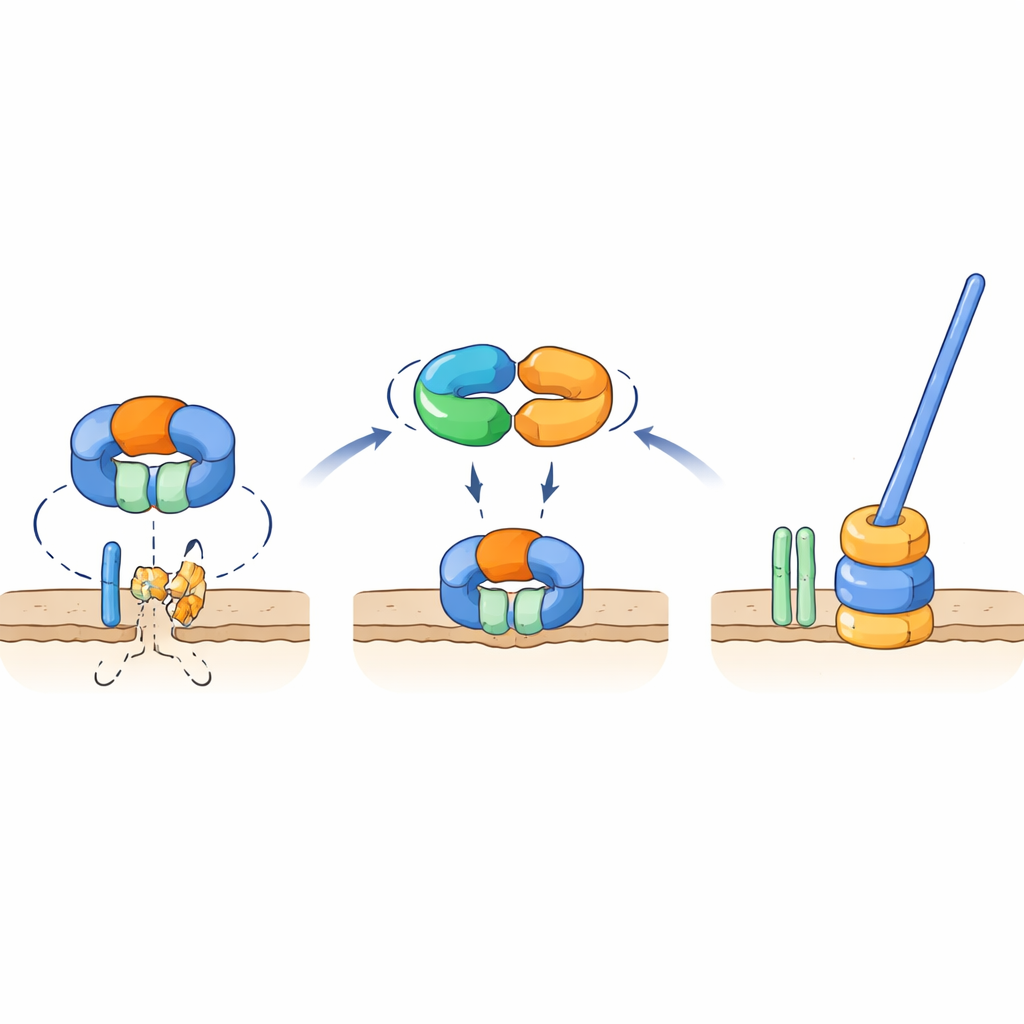
保护关键鞭毛部件的分子开关
遗传学和相互作用测试表明TrxA与PtxA能够物理结合,且EKVEQ基序对这种伙伴关系至关重要。利用细菌双杂交系统,研究者发现TrxA和PtxA还与ClpC相互作用,ClpC是Clp蛋白酶复合体的一个分子伴侣组分——该复合体像一个分子机器,将蛋白质展开并送入桶状“粉碎器”中。为探究这与鞭毛的关系,他们将非运动性的TrxA和PtxA突变体暴露于紫外光并筛选出少数恢复运动性的抑制株。许多抑制株携带ClpC或FliR的突变,FliR是位于鞭毛基部、构成鞭毛输出门的一种膜蛋白。将相同突变引入缺乏TrxA或PtxA的菌株可恢复运动性,而在正常背景下删除fliR则完全阻止鞭毛形成。这些发现支持这样一个模型:在非组装鞭毛的条件下,含ClpC的蛋白酶降解FliR,阻止鞭毛基座的组装。当条件有利于鞭毛形成时,TrxA–PtxA复合体与ClpC结合,抑制其蛋白水解活性,从而使FliR得以积累,输出门随后组装——继而完整鞭毛得以建成。
这项工作对微生物生活和蛋白进化的意义
这项研究揭示了一个精细调控的系统,使细菌能通过保护鞭毛机器中单个易损组件,迅速在休眠孢子与活跃游动体之间切换。它还展示了一种硫氧还蛋白执行并不依赖其常规化学活性的功能:TrxA并非作为氧化还原催化剂,而是通过一个保守的短基序与PtxA形成调控复合体并控制蛋白酶。这类角色转换强调了现有蛋白家族如何在进化过程中被重新利用为新的调控模块,使像A. missouriensis这样的细菌能够仅依靠少数精确的分子互作来协调复杂的生命周期转换。
引用: Kimura, T., Maeda, S., Suzuki, R. et al. Thioredoxin and its partner protein are essential for zoospore flagellar formation in Actinoplanes missouriensis. Commun Biol 9, 532 (2026). https://doi.org/10.1038/s42003-026-09784-8
关键词: 细菌运动, 鞭毛装配, 蛋白质调控, 硫氧还蛋白, 蛋白酶控制