Clear Sky Science · it
La tioredoxina e la sua proteina partner sono essenziali per la formazione dei flagelli negli zoospore di Actinoplanes missouriensis
Come alcuni batteri costruiscono piccole code su richiesta
Molti batteri si muovono usando lunghe code a frusta chiamate flagelli. Per un microbo che vive nel suolo, Actinoplanes missouriensis, costruire queste code nel momento giusto è questione di sopravvivenza: le sue spore devono improvvisamente diventare nuotatrici quando arriva la pioggia o si forma acqua. Questo studio rivela come una coppia di piccole proteine funzioni come un interruttore molecolare on–off per controllare quando quelle code da nuoto vengono costruite.
Dalle spore silenziose ai nuotatori improvvisi
A. missouriensis normalmente cresce come filamenti ramificati nel suolo. In condizioni di aridità forma strutture sferiche dette sporangia sulle punte di questi filamenti. All’interno di ogni sporangio si formano centinaia di spore tonde pronte per la vita esterna. Quando arriva l’acqua, lo sporangio si apre e rilascia le spore nel liquido. Queste spore sviluppano rapidamente flagelli e diventano “zoospore” in grado di nuotare per un breve periodo, aiutandole a disperdersi in nuovi ambienti prima di stabilirsi e ricominciare a crescere. Poiché questa fase di nuoto è breve e strettamente temporizzata, la cellula deve controllare con precisione quando e come i flagelli vengono assemblati.
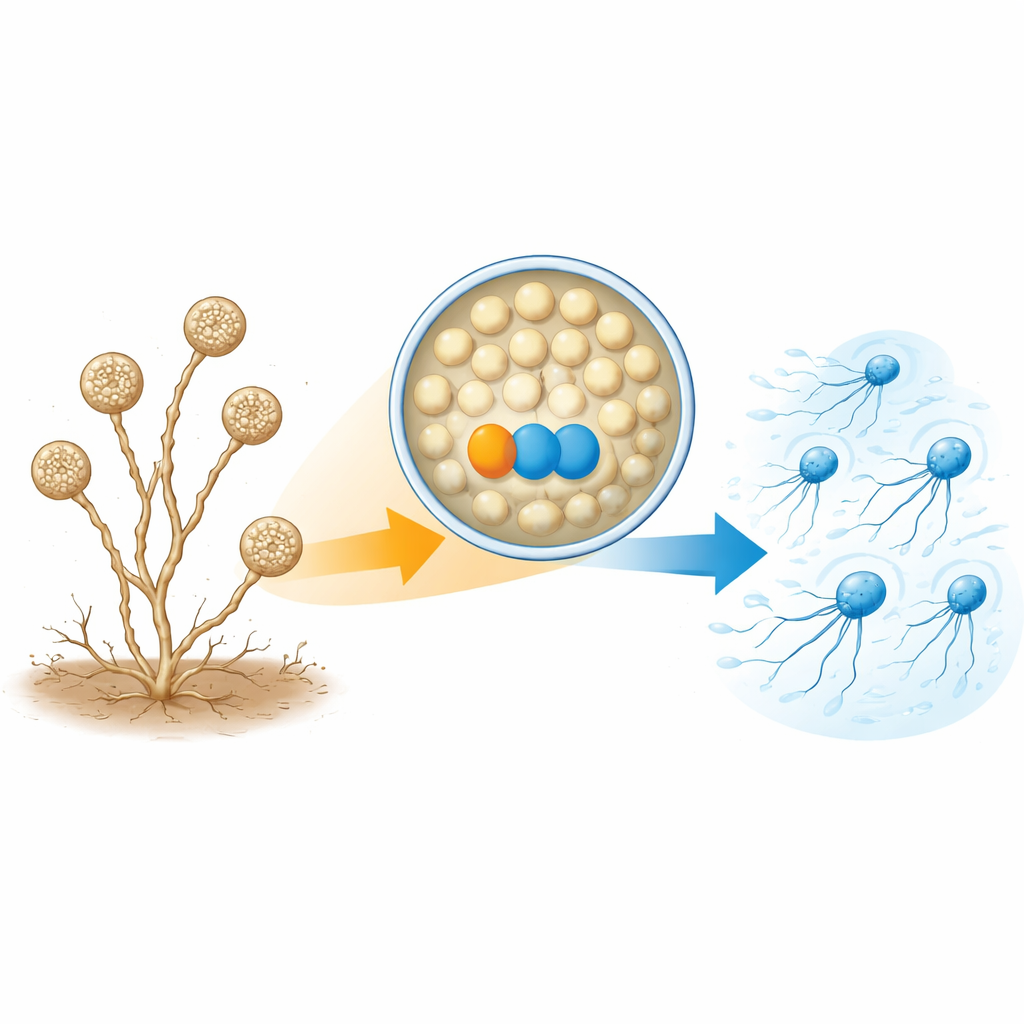
Una coppia di proteine necessaria per costruire le code
Gli autori si sono concentrati su due proteine molto più abbondanti negli zoospore nuotanti che nelle cellule germinanti. Una è una tioredoxina, chiamata TrxA, e l’altra è la sua proteina partner, denominata PtxA. Quando i ricercatori hanno eliminato i geni per TrxA o PtxA, le spore si formarono normalmente all’interno degli sporangia e vennero rilasciate al momento giusto—ma non nuotavano. La microscopia elettronica ha spiegato il motivo: la maggior parte delle spore di questi mutanti mancava semplicemente dei flagelli, o ne aveva solo alcuni corti. Tuttavia i livelli di RNA messaggero dei geni per i flagelli e la quantità di almeno un componente proteico chiave del flagello (FliC) erano essenzialmente normali. Ciò significa che TrxA e PtxA non attivano o disattivano i geni dei flagelli; piuttosto, sono necessari per l’effettivo assemblaggio delle strutture flagellari.
Un ruolo non redox per una classica proteina redox
Le tioredoxine tipicamente operano usando una coppia di residui cisteina per riorganizzare legami disolfuro in altre proteine. Per verificare se questa attività redox classica fosse richiesta in questo caso, il team ha purificato TrxA e ha mostrato in un saggio in provetta che si comporta come una tioredoxina normale: può ridurre un substrato proteico standard. Quando hanno mutato una o entrambe le cisteine chiave, TrxA ha perso questa capacità redox. Sorprendentemente, i batteri che esprimevano queste versioni “redox-dead” di TrxA producevano comunque spore completamente flagellate e motili. Al contrario, sostituire una parte di TrxA con la tioredoxina più simile proveniente da un altro batterio non ha ripristinato la formazione dei flagelli, nonostante la proteina sostitutiva avesse attività tioredossinica normale. Scambiando sistematicamente regioni tra le due proteine, gli autori hanno ristretto la caratteristica cruciale a un breve tratto di cinque amminoacidi in TrxA, con la sequenza EKVEQ, conservato in molte specie di Actinoplanes.
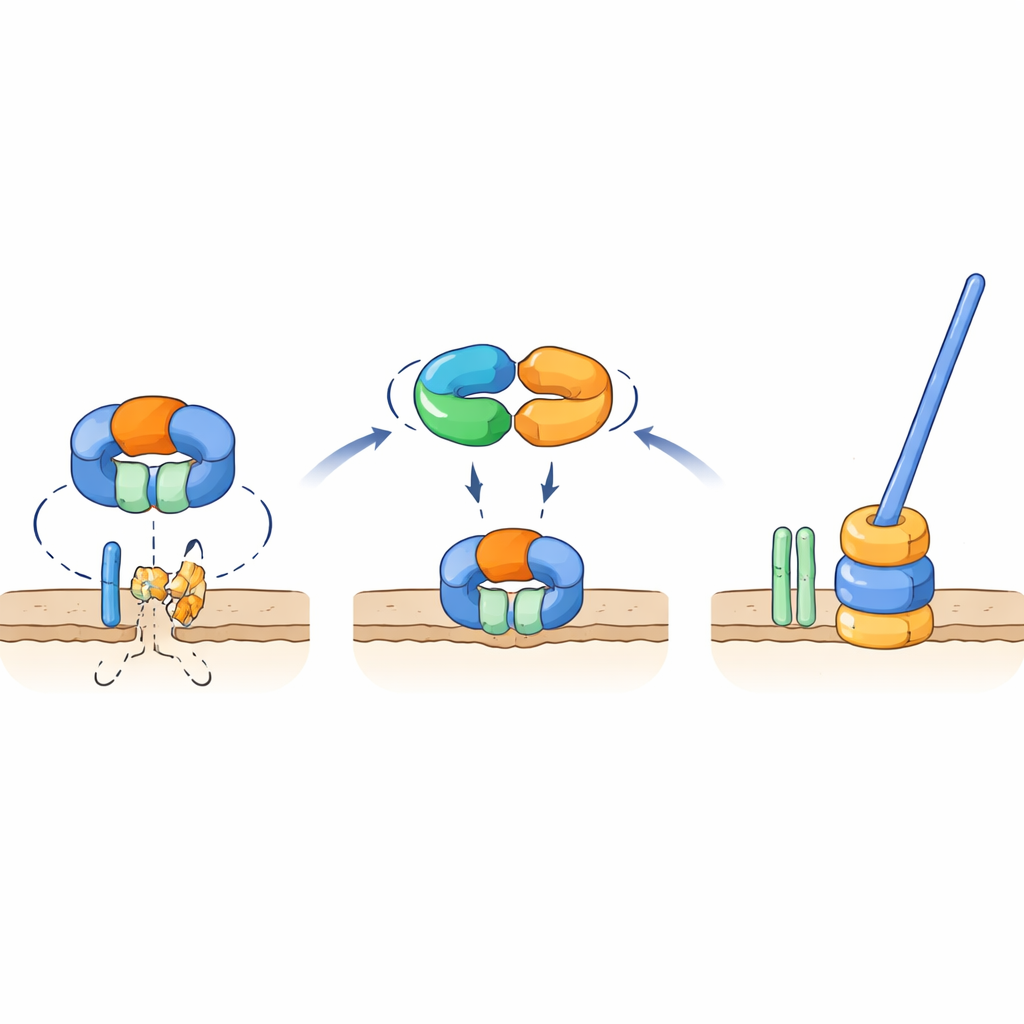
Un interruttore molecolare che protegge una parte chiave del flagello
Test genetici e di interazione hanno mostrato che TrxA e PtxA si legano fisicamente e che il motivo EKVEQ è essenziale per questa associazione. Usando un sistema batterico a due ibridi, i ricercatori hanno scoperto che TrxA e PtxA interagiscono anche con ClpC, un componente chaperone del complesso proteasico Clp—una macchina molecolare che disfa e immette proteine in un “trituratore” a forma di barile. Per capire come questo si ricollega ai flagelli, hanno esposto i mutanti non motili TrxA e PtxA a luce UV e selezionato rari ceppi soppressori le cui spore recuperarono la motilità. Molti di questi soppressori portavano mutazioni in ClpC o in FliR, una proteina di membrana che forma parte del cancello di esportazione flagellare alla base della coda. Introdurre le stesse mutazioni in ceppi privi di TrxA o PtxA ha ripristinato la motilità, mentre la delezione di fliR in un background altrimenti normale aboliva completamente i flagelli. Questi risultati supportano un modello in cui, in condizioni non favorevoli all’assemblaggio dei flagelli, la proteasi contenente ClpC degrada FliR, impedendo la formazione della base flagellare. Quando le condizioni favoriscono la formazione dei flagelli, il complesso TrxA–PtxA si lega a ClpC, attenuandone l’attività proteolitica così che FliR possa accumularsi e il cancello di esportazione—e poi l’intero flagello—possa essere costruito.
Perché questo è importante per la vita microbica e l’evoluzione delle proteine
Questo lavoro rivela un sistema finemente regolato che permette a un batterio di passare rapidamente da una spora dormiente a un nuotatore attivo proteggendo una singola componente vulnerabile della macchina flagellare. Mostra inoltre una tioredoxina che svolge un ruolo indipendente dalla sua chimica abituale: anziché agire come catalizzatore redox, TrxA sfrutta un breve motivo conservato per formare un complesso regolatorio con PtxA e controllare una proteasi. Questo tipo di cambio di ruolo evidenzia come famiglie proteiche esistenti possano essere riutilizzate durante l’evoluzione in nuovi moduli regolatori, permettendo a batteri come A. missouriensis di coordinare transizioni complesse del ciclo vitale con poche, ben posizionate interazioni molecolari.
Citazione: Kimura, T., Maeda, S., Suzuki, R. et al. Thioredoxin and its partner protein are essential for zoospore flagellar formation in Actinoplanes missouriensis. Commun Biol 9, 532 (2026). https://doi.org/10.1038/s42003-026-09784-8
Parole chiave: moto batterico, assemblaggio dei flagelli, regolazione proteica, tioredoxina, controllo della proteasi