Clear Sky Science · de
Thioredoxin und sein Partnerprotein sind für die Bildung von Zoosporen-Flagellen in Actinoplanes missouriensis unerlässlich
Wie manche Bakterien kleine Schwänze auf Abruf bauen
Viele Bakterien bewegen sich mit langen, peitschenartigen Anhängseln, den Flagellen. Für einen im Boden lebenden Mikroorganismus namens Actinoplanes missouriensis ist das exakte Timing beim Bau dieser Anhängsel überlebenswichtig: Seine Sporen müssen beim Auftreten von Regen oder Wasser plötzlich zu Schwimmern werden. Diese Studie zeigt, wie ein Paar kleiner Proteine wie ein molekularer Ein-/Ausschalter wirkt und kontrolliert, wann diese Schwänzchen gebaut werden.
Von ruhenden Sporen zu plötzlichen Schwimmern
A. missouriensis wächst normalerweise als verzweigte Filamente im Boden. Unter trockenen Bedingungen bildet es an den Spitzen dieser Filamente runde Strukturen, sogenannte Sporangien. In jedem Sporangium entstehen Hunderte runder Sporen, die sich auf das Leben außerhalb vorbereiten. Wenn Wasser eintrifft, öffnet sich das Sporangium und setzt die Sporen in die Flüssigkeit frei. Diese Sporen treiben schnell Flagellen aus und werden zu „Zoosporen“, die für kurze Zeit schwimmen können und so helfen, neue Lebensräume zu erreichen, bevor sie sich wieder niederlassen und erneut wachsen. Weil diese Schwimmphase kurz und streng getimt ist, muss die Zelle genau kontrollieren, wann und wie Flagellen zusammengesetzt werden.
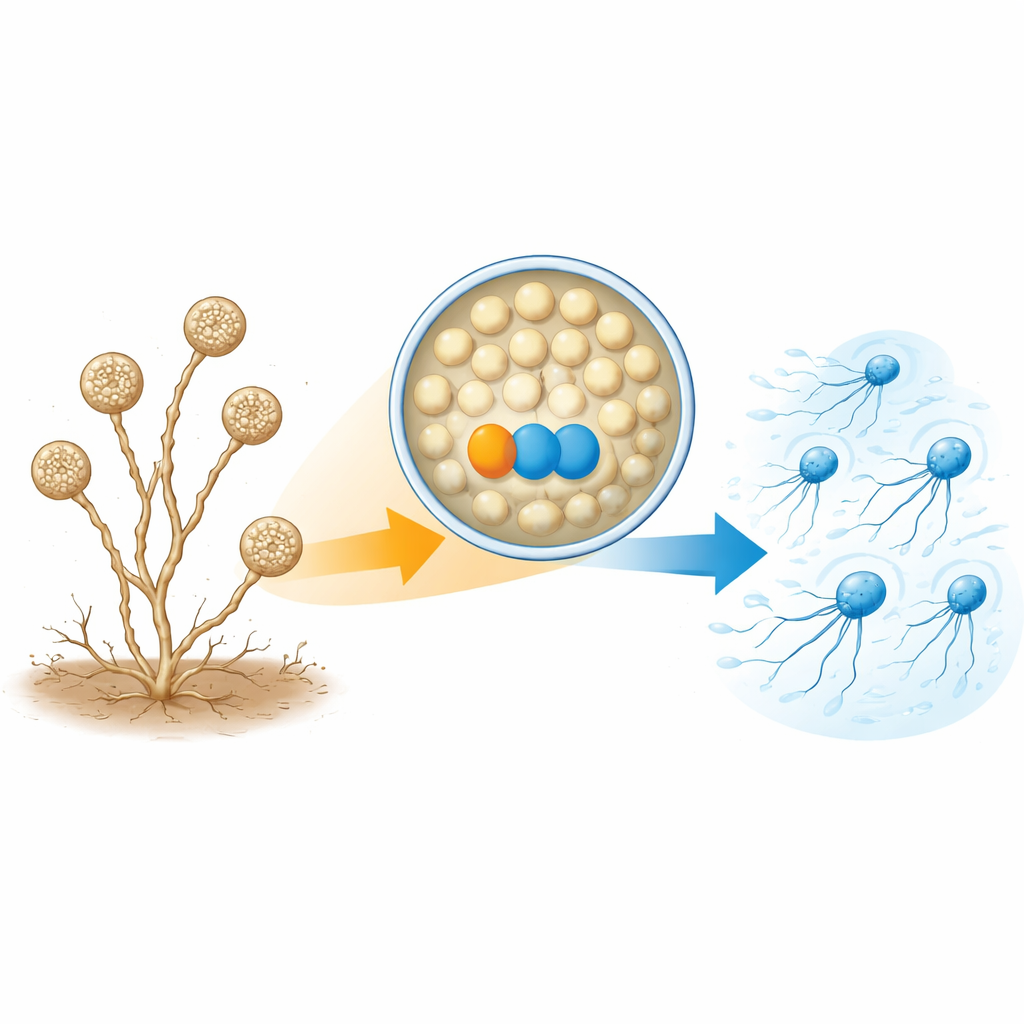
Ein Proteinpaar, das zum Bau der Schwänze benötigt wird
Die Autor:innen konzentrierten sich auf zwei Proteine, die in schwimmenden Zoosporen deutlich stärker vorhanden sind als in keimenden Zellen. Das eine ist ein Thioredoxin namens TrxA, das andere sein Partnerprotein PtxA. Wenn die Forschenden die Gene für TrxA oder PtxA entfernten, bildeten die Sporen im Sporangium weiterhin normal und wurden wie erwartet freigesetzt – sie schwammen jedoch nicht. Elektronenmikroskopie zeigte den Grund: Die meisten Sporen der Mutanten hatten gar keine Flagellen oder nur wenige kurze. Die mRNA-Level der Flagellengene und die Menge zumindest eines wichtigen Flagellen-Bausteinproteins (FliC) waren weitgehend normal. Das bedeutet, TrxA und PtxA schalten Flagellengene nicht an oder aus; sie sind vielmehr für den tatsächlichen Aufbau der Flagellenstrukturen erforderlich.
Eine nicht-redoxmäßige Aufgabe für ein klassisches Redoxprotein
Thioredoxine arbeiten typischerweise, indem sie ein Paar Cystein-Aminosäuren verwenden, um Disulfidbrücken in anderen Proteinen zu verschieben. Um zu prüfen, ob diese klassische Redoxaktivität hier nötig ist, reinigten die Forschenden TrxA und zeigten in einem Reagenzglas-Assay, dass es sich wie ein normales Thioredoxin verhält: Es kann ein Standardsubstrat reduzieren. Wenn sie eines oder beide Schlüsselcysteine veränderten, verlor TrxA diese Redoxfähigkeit. Überraschenderweise produzierten Bakterien mit diesen „redox-inaktiven“ Versionen von TrxA dennoch voll flagellierte, bewegliche Sporen. Im Gegensatz dazu konnte der Tausch eines Teils von TrxA durch das nächstähnliche Thioredoxin eines anderen Bakteriums die Flagellenbildung nicht retten, obwohl das Ersatzprotein normale Thioredoxin-Aktivität zeigte. Durch systematischen Austausch von Regionen zwischen den beiden Proteinen schränkten die Autor:innen das entscheidende Merkmal auf einen kurzen Fünf-Aminosäure-Abschnitt in TrxA mit der Sequenz EKVEQ ein, der in vielen Arten von Actinoplanes konserviert ist.
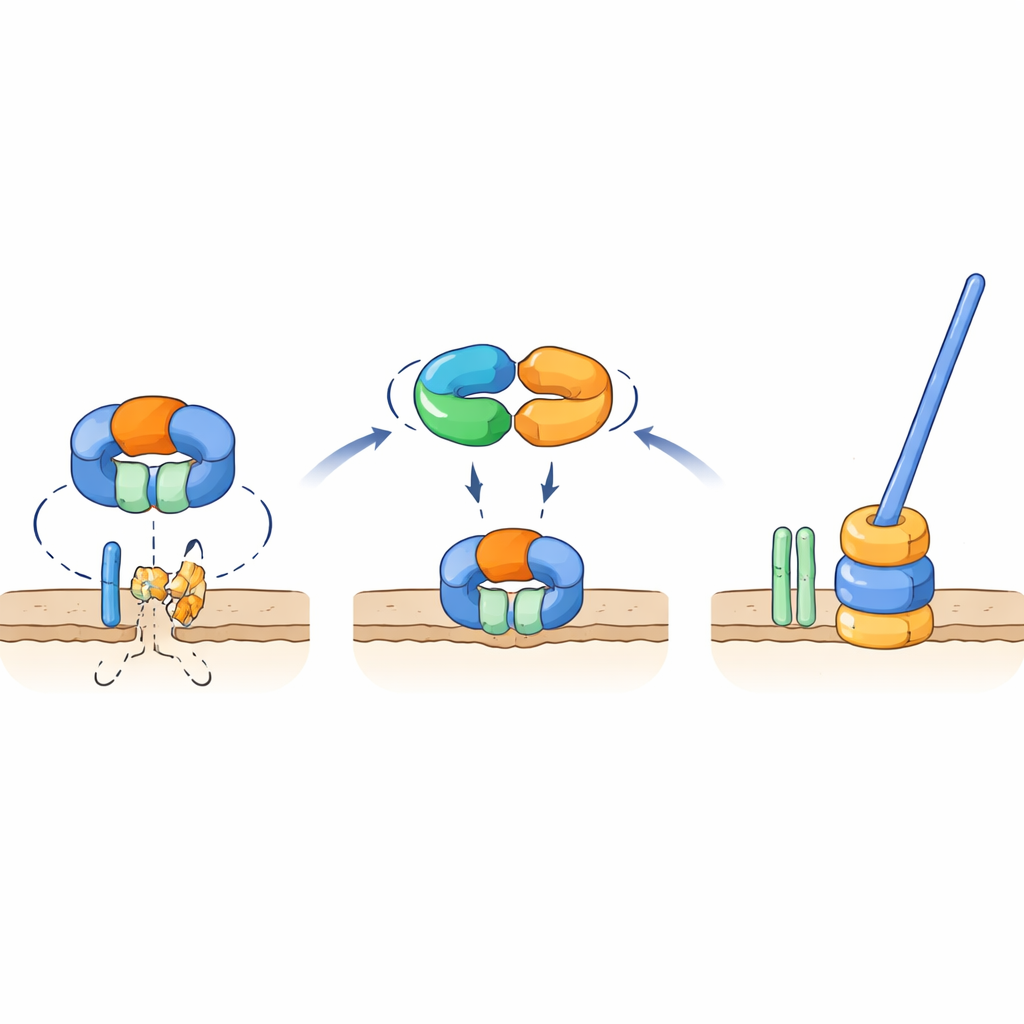
Ein molekularer Schalter, der ein wichtiges Flagellenteil schützt
Genetische und Wechselwirkungstests zeigten, dass TrxA und PtxA physikalisch aneinander binden und dass das EKVEQ-Motiv für diese Partnerschaft essentiell ist. Mithilfe eines bakteriellen Zwei-Hybrid-Systems fanden die Forschenden heraus, dass TrxA und PtxA auch mit ClpC interagieren, einer Chaperon-Komponente des Clp-Protease-Komplexes – einer molekularen Maschine, die Proteine entfaltet und in ein tonnenförmiges „Schredder“-System einspeist. Um zu untersuchen, wie das mit den Flagellen zusammenhängt, setzten sie die nicht-mobilen TrxA- und PtxA-Mutanten UV-Licht aus und selektierten seltene Suppressorstämme, deren Sporen die Beweglichkeit zurückerlangten. Viele dieser Suppressoren trugen Mutationen in ClpC oder in FliR, einem Membranprotein, das Teil des Flagellen-Exporttors an der Basis des Schwanzes ist. Die Einführung derselben Mutationen in Stämmen ohne TrxA oder PtxA stellte die Beweglichkeit wieder her, und die Deletion von fliR in einem ansonsten normalen Hintergrund beseitigte die Flagellen vollständig. Diese Befunde stützen ein Modell, in dem das ClpC-haltige Protease-System unter nicht-flagellierenden Bedingungen FliR abbaut und so den Aufbau der Flagellenbasis verhindert. Wenn die Bedingungen den Flagellenaufbau begünstigen, bindet der TrxA–PtxA-Komplex an ClpC und dämpft dessen proteolytische Aktivität, sodass FliR akkumulieren kann und das Exporttor – und anschließend die komplette Flagelle – aufgebaut werden kann.
Warum das für mikrobielles Leben und die Protein-Evolution wichtig ist
Diese Arbeit offenbart ein fein ausbalanciertes System, das es einem Bakterium erlaubt, rasch zwischen einer ruhenden Spore und einem aktiven Schwimmer zu wechseln, indem eine einzelne verwundbare Komponente der Flagellenmaschine geschützt wird. Sie zeigt außerdem, dass ein Thioredoxin eine Funktion ausübt, die nicht von seiner üblichen Chemie abhängt: Statt als Redox-Katalysator zu wirken, nutzt TrxA ein kurzes, konserviertes Motiv, um einen regulatorischen Komplex mit PtxA zu bilden und eine Protease zu kontrollieren. Solche Rollenwechsel veranschaulichen, wie bestehende Protein-Familien evolutionär in neue Regulationsmodule umgewidmet werden können, sodass Bakterien wie A. missouriensis komplexe Lebenszyklus-Übergänge mit nur einer Handvoll gezielt platzierter molekularer Interaktionen koordinieren können.
Zitation: Kimura, T., Maeda, S., Suzuki, R. et al. Thioredoxin and its partner protein are essential for zoospore flagellar formation in Actinoplanes missouriensis. Commun Biol 9, 532 (2026). https://doi.org/10.1038/s42003-026-09784-8
Schlüsselwörter: bakterielle Motilität, Flagellenaufbau, Proteinregulation, Thioredoxin, Proteasekontrolle