Clear Sky Science · ru
Тиоредоксин и его партнерская белковая пара необходимы для формирования жгутиков зооспор у Actinoplanes missouriensis
Как некоторые бактерии по требованию собирают крошечные хвостики
Многие бактерии передвигаются с помощью длинных, хлыстовидных структур, называемых жгутиками. Для почвенной микробной формы жизни Actinoplanes missouriensis сборка этих хвостиков в нужный момент — вопрос выживания: ее споры должны внезапно превратиться в плавающие клетки, когда появляется дождь или влага. В этой статье показано, как пара малых белков действует как молекулярный выключатель, контролируя, когда эти плавающие хвостики собираются.
От молчаливых спор к внезапным пловцам
A. missouriensis обычно растет в виде разветвленных нитей в почве. В сухих условиях на кончиках этих нитей образуются круглые структуры — спорангии. Внутри каждого спорангия формируются сотни округлых спор, подготовленных к жизни вне клетки. Когда приходит вода, спорангий раскрывается и споры попадают в жидкость. Эти споры быстро формируют жгутики и становятся «зооспорами», которые некоторое время плавают, помогая им распространиться в новые места, прежде чем осесть и снова начать рост. Поскольку фаза плавания коротка и строго синхронизирована, клетке нужно точно контролировать, когда и как собираются жгутики.
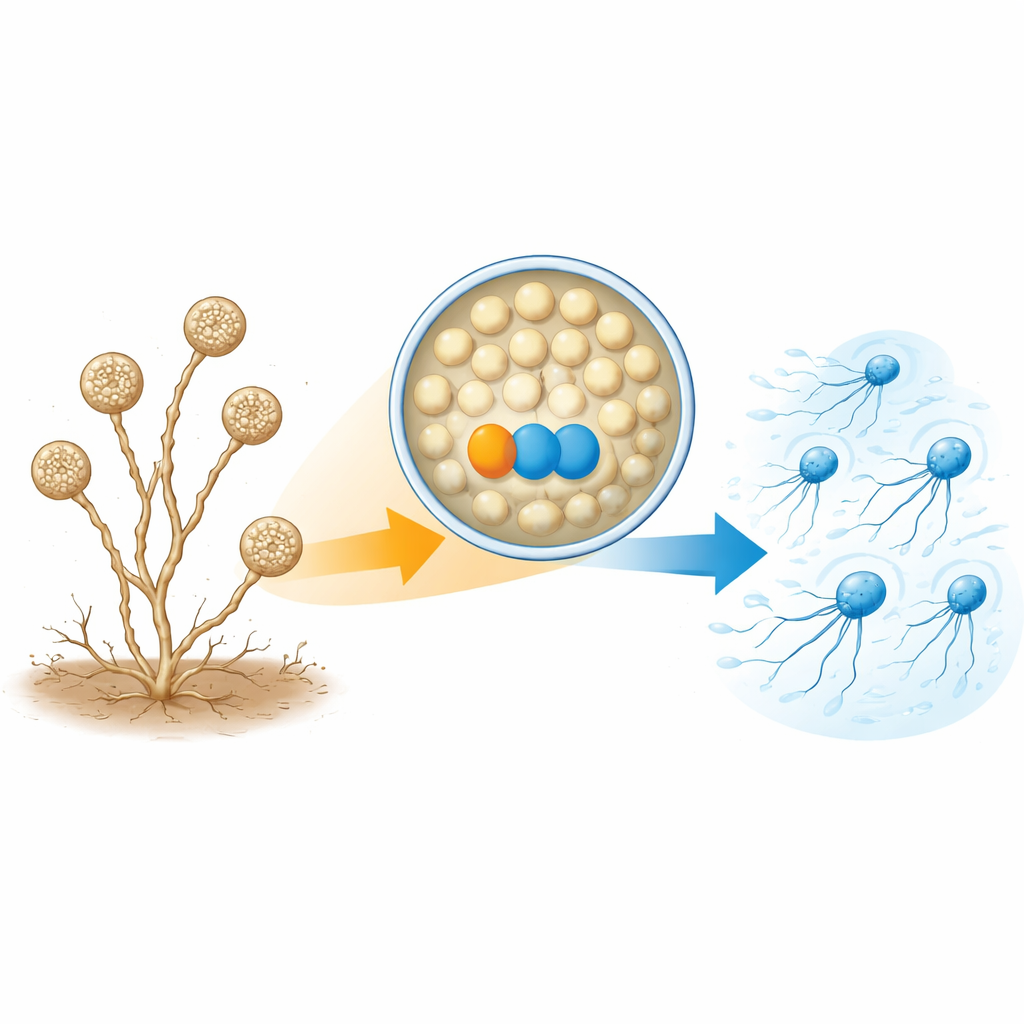
Парa белков, необходимая для строительства хвостиков
Авторы сосредоточили внимание на двух белках, которые значительно более многочисленны в плавающих зооспорах, чем в прорастающих клетках. Один из них — тиоредоксин, названный TrxA, а другой — его партнерский белок PtxA. При удалении генов TrxA или PtxA споры нормально формировались внутри спорангиев и своевременно высвобождались — но не плавали. Электронная микроскопия показала причину: у большинства спор этих мутантов либо полностью отсутствовали жгутики, либо оставались лишь несколько коротких. При этом уровни мРНК для генов жгутика и количество по крайней мере одного ключевого строительного белка жгутика (FliC) оставались по существу нормальными. Это означает, что TrxA и PtxA не включают и не выключают синтез генов жгутика; вместо этого они необходимы для самой сборки жгутиковых структур.
Нередоксная роль классического редокс-белка
Тиоредоксины обычно работают, используя пару цистеинов для перестановки дисульфидных связей в других белках. Чтобы проверить, нужна ли здесь такая классическая редокс-активность, команда очистила TrxA и показала в пробирочном тесте, что он ведет себя как обычный тиоредоксин: может восстановливать стандартный белковый субстрат. Когда исследователи заменили одну или обе ключевые цистеины, TrxA потерял эту редокс-способность. Удивительно, но бактерии, несущие эти «редокс-мертвые» версии TrxA, по-прежнему давали полностью жгутиконосные и подвижные споры. Напротив, замена части TrxA ближайшим похожим тиоредоксином из другой бактерии не восстановила формирование жгутиков, хотя заменяющий белок имел нормальную тиоредоксиновую активность. Путем систематического обмена регионов между двумя белками авторы сузили критическую особенность до короткого участка из пяти аминокислот в TrxA с последовательностью EKVEQ, который сохранен во многих видах Actinoplanes.
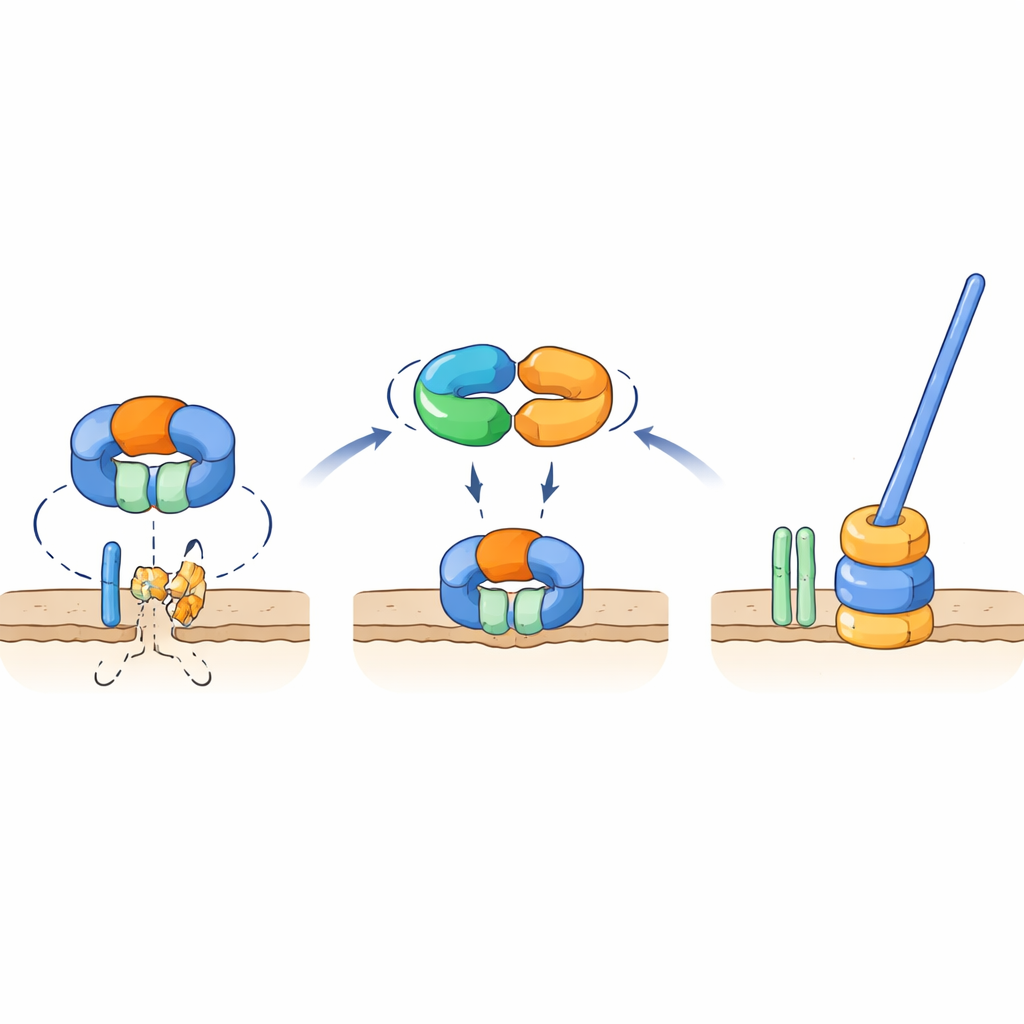
Молекулярный переключатель, который защищает ключевую часть жгутика
Генетические и взаимодействующие тесты показали, что TrxA и PtxA физически связываются друг с другом, и что мотив EKVEQ необходим для этого партнерства. С помощью бактериальной двухгибридной системы исследователи обнаружили, что TrxA и PtxA также взаимодействуют с ClpC — шаперонной компонентой комплекса протеаз Clp, молекулярной машины, которая разворачивает белки и направляет их в бочкообразный «измельчитель». Чтобы выяснить, как это связано с жгутиком, они подвергли немотильные мутанты TrxA и PtxA УФ-облучению и отобрали редкие супрессорные штаммы, чьи споры вновь обрели подвижность. Многие из этих супрессоров несли мутации в ClpC или в FliR, мембранном белке, который формирует часть экспортных ворот жгутика у его основания. Введение тех же мутаций обратно в штаммы, лишенные TrxA или PtxA, восстановило подвижность, а удаление fliR в иначе нормальном фоне полностью устраняло жгутики. Эти результаты поддерживают модель, в которой при условиях, не благоприятствующих формированию жгутика, протеаза с участием ClpC разрушает FliR, препятствуя сборке основания жгутика. Когда условия становятся благоприятными, комплекс TrxA–PtxA связывается с ClpC, ослабляя его протеолитическую активность, что позволяет накопиться FliR и образоваться экспортным воротам — а затем и полному жгутику.
Почему это важно для микробной жизни и эволюции белков
Эта работа выявляет тонко настроенную систему, которая позволяет бактерии быстро переключаться между спящими спорами и активными пловцами, защищая единственный уязвимый компонент жгутикового аппарата. Она также показывает, что тиоредоксин выполняет роль, не зависящую от его обычной химии: вместо редокс-катализа TrxA использует короткий сохраненный мотив для формирования регуляторного комплекса с PtxA и контроля протеазы. Такой переход функции иллюстрирует, как существующие семейства белков могут перепрофилироваться в ходе эволюции в новые регуляторные модули, позволяя бактериям вроде A. missouriensis согласовывать сложные переходы в жизненном цикле с помощью всего нескольких точных молекулярных взаимодействий.
Цитирование: Kimura, T., Maeda, S., Suzuki, R. et al. Thioredoxin and its partner protein are essential for zoospore flagellar formation in Actinoplanes missouriensis. Commun Biol 9, 532 (2026). https://doi.org/10.1038/s42003-026-09784-8
Ключевые слова: бактериальная подвижность, сборка жгутика, регуляция белков, тиоредоксин, контроль протеаз