Clear Sky Science · pl
Tioredoksyna i jej białkowy partner są niezbędne do formowania witki zoospor w Actinoplanes missouriensis
Jak niektóre bakterie budują maleńkie ogony na żądanie
Wiele bakterii porusza się przy pomocy długich, biczowatych struktur zwanych witekami. Dla mikroorganizmu żyjącego w glebie, Actinoplanes missouriensis, budowa tych ogonów we właściwym momencie to kwestia przetrwania: jego zarodniki muszą nagle stać się pływakami, gdy pojawi się deszcz lub woda. Artykuł ten ujawnia, jak para małych białek działa jak molekularny włącznik-wyłącznik kontrolujący, kiedy te pływackie witki są wytwarzane.
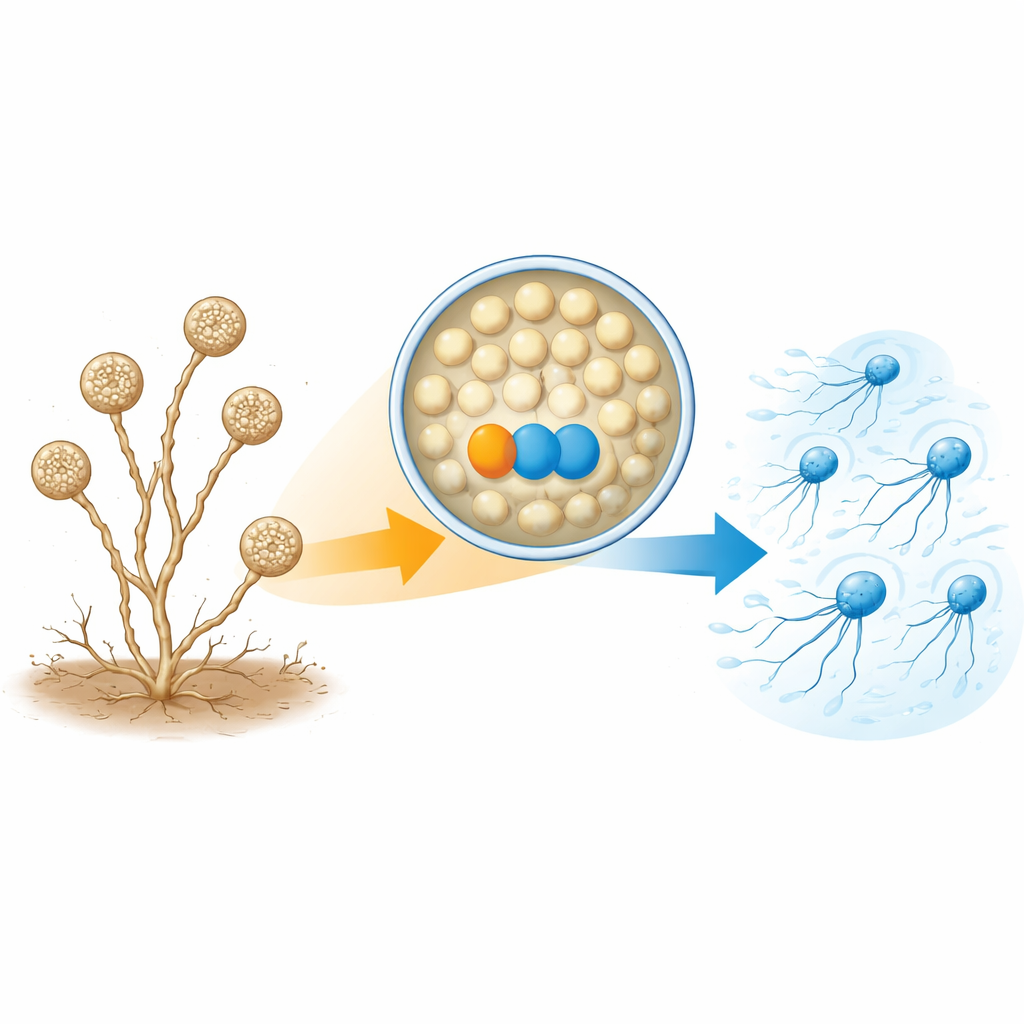
Z cichych zarodników do nagłych pływaków
A. missouriensis zazwyczaj rośnie jako rozgałęzione strzępki w glebie. W suchych warunkach tworzy na końcach tych strzępek okrągłe struktury zwane sporangami. W każdym sporangium powstają setki okrągłych zarodników przygotowanych do życia poza strukturą. Gdy pojawia się woda, sporangium otwiera się i uwalnia zarodniki do cieczy. Zarodniki te szybko wyrastają witki i stają się „zoosporami”, które przez krótki czas potrafią pływać, co pomaga im rozprzestrzeniać się w nowe miejsca, zanim osiedlą się i znów zaczną rosnąć. Ponieważ faza pływania jest krótka i ściśle zsynchronizowana, komórka musi starannie kontrolować dokładny moment i sposób składania witek.
Para białek niezbędna do budowy ogonów
Autorzy skupili się na dwóch białkach, które są znacznie obficiej obecne w pływających zoosporach niż w kiełkujących komórkach. Jedno to tioredoksyna, nazwana TrxA, a drugie to jej partner, oznaczony jako PtxA. Gdy badacze usunęli geny kodujące albo TrxA, albo PtxA, zarodniki nadal formowały się normalnie w sporangiach i uwalniane były na sygnał — jednak nie pływały. Mikroskopia elektronowa wyjaśniła przyczynę: większość zarodników tych mutantów po prostu nie miała witek albo miała tylko kilka krótkich. Mimo to poziomy mRNA dla genów witek oraz ilość przynajmniej jednego kluczowego białka budulcowego witek (FliC) były w zasadzie normalne. Oznacza to, że TrxA i PtxA nie włączają ani nie wyłączają genów witek; zamiast tego są potrzebne do samego montażu struktur witek.
Nie-redoksowa rola klasycznego białka redoks
Tioredoksyny zwykle działają, wykorzystując parę reszt cysteiny do przebudowy mostków disiarczkowych w innych białkach. Aby sprawdzić, czy ta klasyczna aktywność redoks jest potrzebna tutaj, zespół oczyścił TrxA i pokazał w teście probówkowym, że zachowuje się jak typowa tioredoksyna: może redukować standardowe białkowe substraty. Gdy zmieniono jedną lub obie kluczowe cysteiny, TrxA stracił tę aktywność redoksową. Zaskakująco, bakterie niosące takie „martwe redoksowo” wersje TrxA nadal wytwarzały w pełni uzbrojone w witki, ruchome zarodniki. Natomiast zamiana części TrxA na najbliższą podobną tioredoksynę z innej bakterii nie przywróciła formowania witek, mimo że zastępcze białko miało normalną aktywność tioredoksyny. Poprzez systematyczną wymianę regionów między dwoma białkami autorzy zawęzili kluczową cechę do krótkiego pięcioaminokwasowego fragmentu w TrxA o sekwencji EKVEQ, który jest konserwowany w wielu gatunkach Actinoplanes.
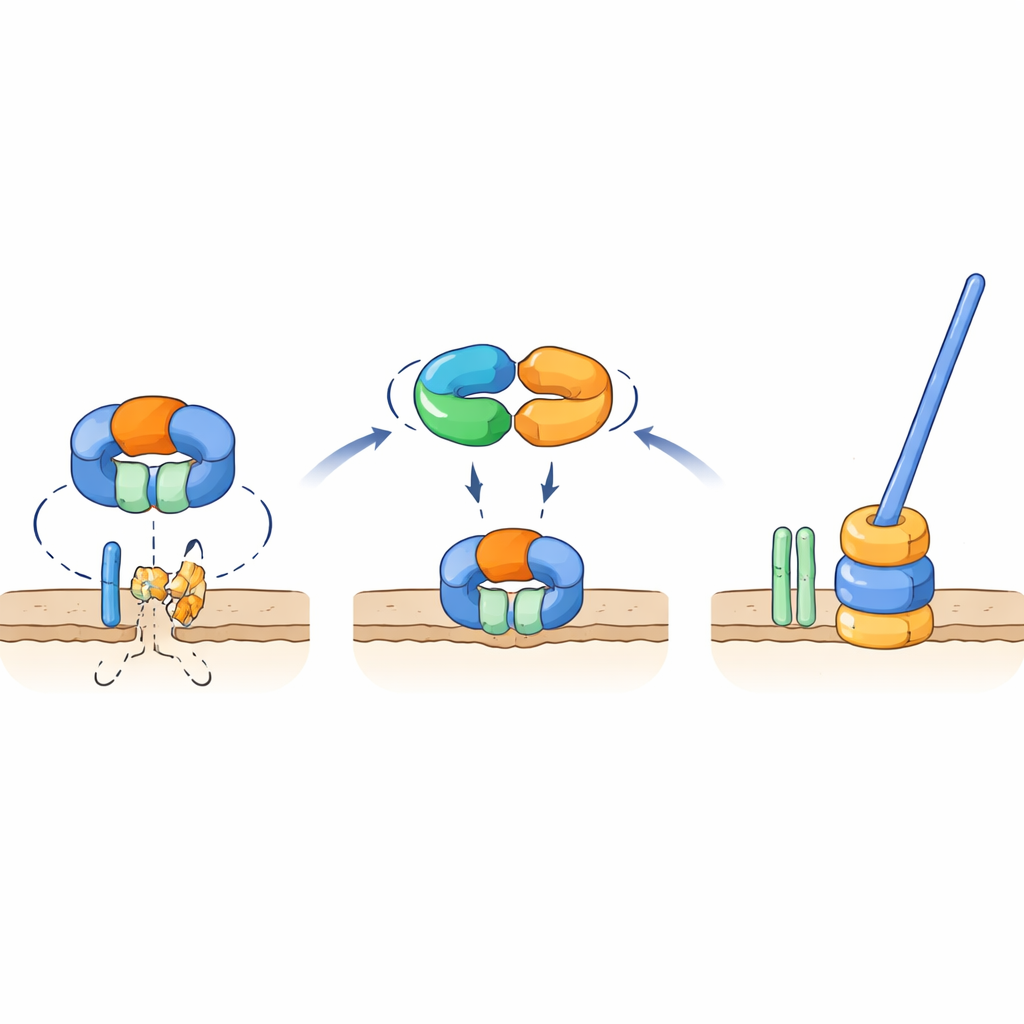
Molekularny przełącznik, który chroni kluczowy element witek
Badania genetyczne i testy interakcji wykazały, że TrxA i PtxA fizycznie wiążą się ze sobą, a motyw EKVEQ jest niezbędny do tej współpracy. Przy użyciu bakteryjnego systemu dwóch hybryd badacze odkryli, że TrxA i PtxA również wchodzą w interakcje z ClpC, chaperonem będącym częścią kompleksu proteazy Clp — molekularnej maszyny, która rozwija białka i podaje je do cylindrycznego „niszczarza”. Aby sprawdzić, jak to odnosi się do witek, poddali nie-ruchome mutanty TrxA i PtxA działaniu promieni UV i wyizolowali rzadkie supresorowe szczepy, których zarodniki odzyskały ruch. Wiele z tych supresorów miało mutacje w ClpC lub w FliR, białku błonowym tworzącym część bramki eksportowej w podstawie witek. Wprowadzenie tych samych mutacji z powrotem do szczepów pozbawionych TrxA lub PtxA przywracało ruch, a usunięcie fliR w z pozoru normalnym tle całkowicie znosiło formowanie witek. Wyniki te wspierają model, w którym w warunkach niedogodnych dla formowania witek, proteaza z ClpC degraduje FliR, zapobiegając zmontowaniu podstawy witek. Gdy warunki sprzyjają budowie witek, kompleks TrxA–PtxA wiąże się z ClpC, tłumiąc jego aktywność proteolityczną, dzięki czemu FliR może się kumulować, bramka eksportowa — a następnie cała witka — mogą zostać zbudowane.
Dlaczego to ma znaczenie dla życia drobnoustrojów i ewolucji białek
Praca ta ujawnia precyzyjnie wyregulowany system, który pozwala bakterii szybko przełączać się między uśpionym zarodnikiem a aktywnym pływakiem, chroniąc pojedynczy wrażliwy element maszyny witek. Pokazuje też tioredoksynę wykonującą zadanie niezależne od jej zwykłej chemii: zamiast działać jako katalizator redoksowy, TrxA wykorzystuje krótki konserwowany motyw do tworzenia kompleksu regulacyjnego z PtxA i kontroli proteazy. Taka zmiana roli podkreśla, jak istniejące rodziny białek mogą zostać zaadaptowane w toku ewolucji do nowych modułów regulacyjnych, umożliwiając bakteriom takim jak A. missouriensis koordynowanie złożonych przejść cyklu życiowego przy użyciu zaledwie kilku precyzyjnie rozmieszczonych interakcji molekularnych.
Cytowanie: Kimura, T., Maeda, S., Suzuki, R. et al. Thioredoxin and its partner protein are essential for zoospore flagellar formation in Actinoplanes missouriensis. Commun Biol 9, 532 (2026). https://doi.org/10.1038/s42003-026-09784-8
Słowa kluczowe: ruch bakteryjny, montaż witek, regulacja białek, tioredoksyna, kontrola proteazy