Clear Sky Science · nl
Thioredoxine en zijn partner-eiwit zijn essentieel voor de vorming van zoospore-flagellen in Actinoplanes missouriensis
Hoe sommige bacteriën op verzoek piepkleine staarten bouwen
Veel bacteriën bewegen zich met lange, zweepachtige staarten die flagellen worden genoemd. Voor een bodembewonende microbe genaamd Actinoplanes missouriensis is het bouwen van die staarten op precies het juiste moment van levensbelang: zijn sporen moeten plotseling zwemvaardig worden wanneer regen of water verschijnt. Dit artikel onthult hoe een paar kleine eiwitten werken als een moleculaire aan-uit-schakelaar om te bepalen wanneer die zwemstaarten worden gebouwd.
Van stille sporen naar plotselinge zwemmers
A. missouriensis groeit normaal gesproken als vertakte draden in de bodem. Onder droge omstandigheden maakt het ronde structuren, sporangia, aan het uiteinde van die draden. In elk sporangium vormen honderden ronde sporen en bereiden ze zich voor op het leven buiten. Als er water komt, opent het sporangium zich en worden de sporen in de vloeistof vrijgelaten. Deze sporen ontwikkelen snel flagellen en worden “zoosporen” die korte tijd kunnen zwemmen, wat hen helpt zich naar nieuwe plaatsen te verspreiden voordat ze zich opnieuw vestigen om te groeien. Omdat deze zwemfase kort en nauwkeurig getimed is, moet de cel precies regelen wanneer en hoe flagellen worden geassembleerd.
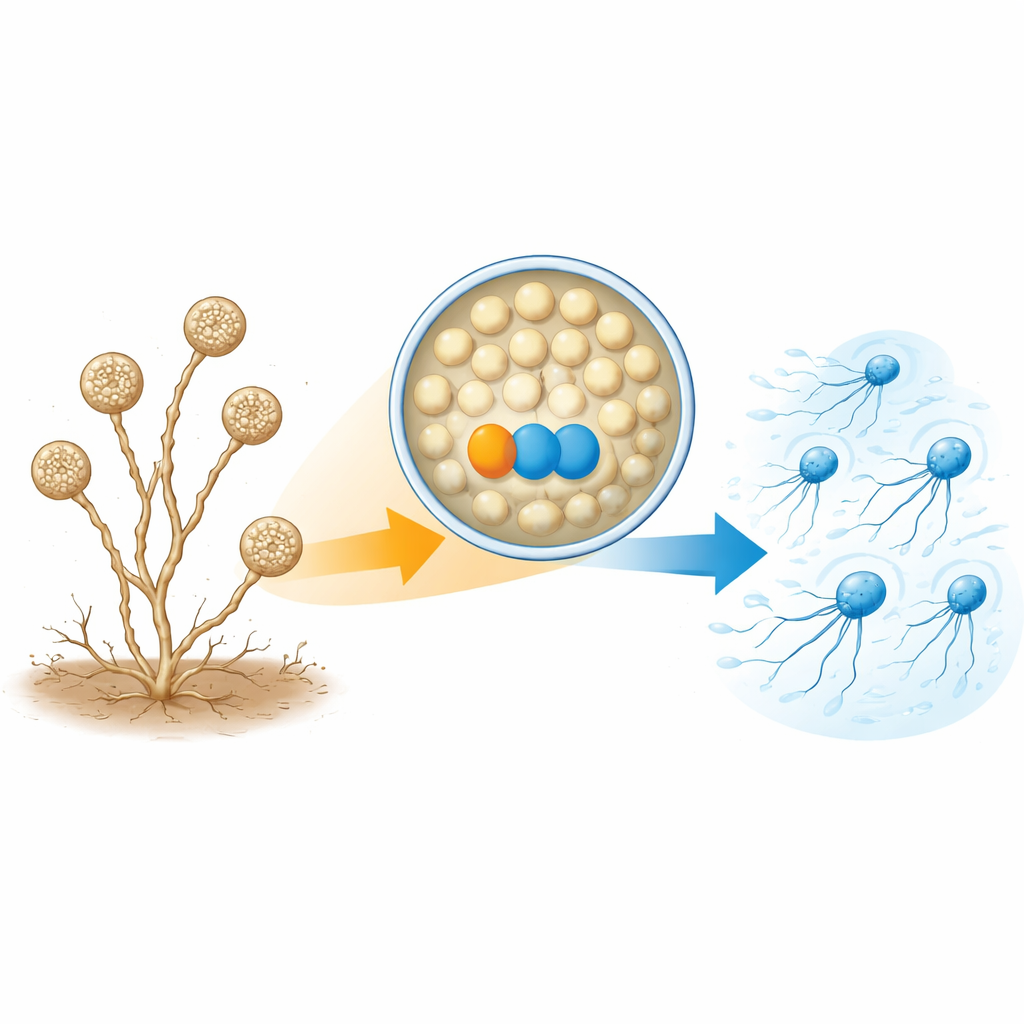
Een eiwitpaar dat nodig is om staarten te bouwen
De auteurs richtten zich op twee eiwitten die veel meer aanwezig zijn in zwemmende zoosporen dan in kiemende cellen. De ene is een thioredoxine, genoemd TrxA, en de andere is het partner-eiwit PtxA. Toen de onderzoekers de genen voor óf TrxA óf PtxA verwijderden, vormden de sporen nog steeds normaal in de sporangia en werden ze op het juiste moment vrijgegeven — maar ze konden niet zwemmen. Elektronenmicroscopie toonde waarom: de meeste sporen van deze mutanten misten gewoon flagellen, of hadden slechts een paar korte exemplaren. Toch waren de messenger-RNA-niveaus van de flagellagenen en de hoeveelheid van ten minste één sleuteleiwit voor de flagelconstructie (FliC) in wezen normaal. Dit betekent dat TrxA en PtxA de flagellagenen niet aan- of uitzetten; in plaats daarvan zijn ze vereist voor de daadwerkelijke assemblage van de flagellairstructuren.
Een niet-redoxtaak voor een klassiek redox-eiwit
Thioredoxines werken meestal door een paar cysteïne-aminozuren te gebruiken om disulfidebindingen in andere eiwitten te herschikken. Om te testen of deze klassieke redoxactiviteit hier nodig was, zuiverde het team TrxA en toonde in een buisproef aan dat het zich gedraagt als een normale thioredoxine: het kan een standaard eiwitsubstraat reduceren. Toen ze één of beide sleutelcysteïnes wijzigden, verloor TrxA deze redoxcapaciteit. Verrassend genoeg produceerden bacteriën met deze “redox-dode” versies van TrxA nog steeds volledig geflagelleerde, beweeglijke sporen. Daarentegen herstelde het vervangen van een deel van TrxA door de meest verwante thioredoxine uit een andere bacterie de flagelvorming niet, hoewel het vervangende eiwit normale thioredoxine-activiteit had. Door systematisch regio’s tussen de twee eiwitten uit te wisselen, beperkten de auteurs de cruciale eigenschap tot een korte stretch van vijf aminozuren in TrxA, met de sequentie EKVEQ, die conserved is in veel soorten Actinoplanes.
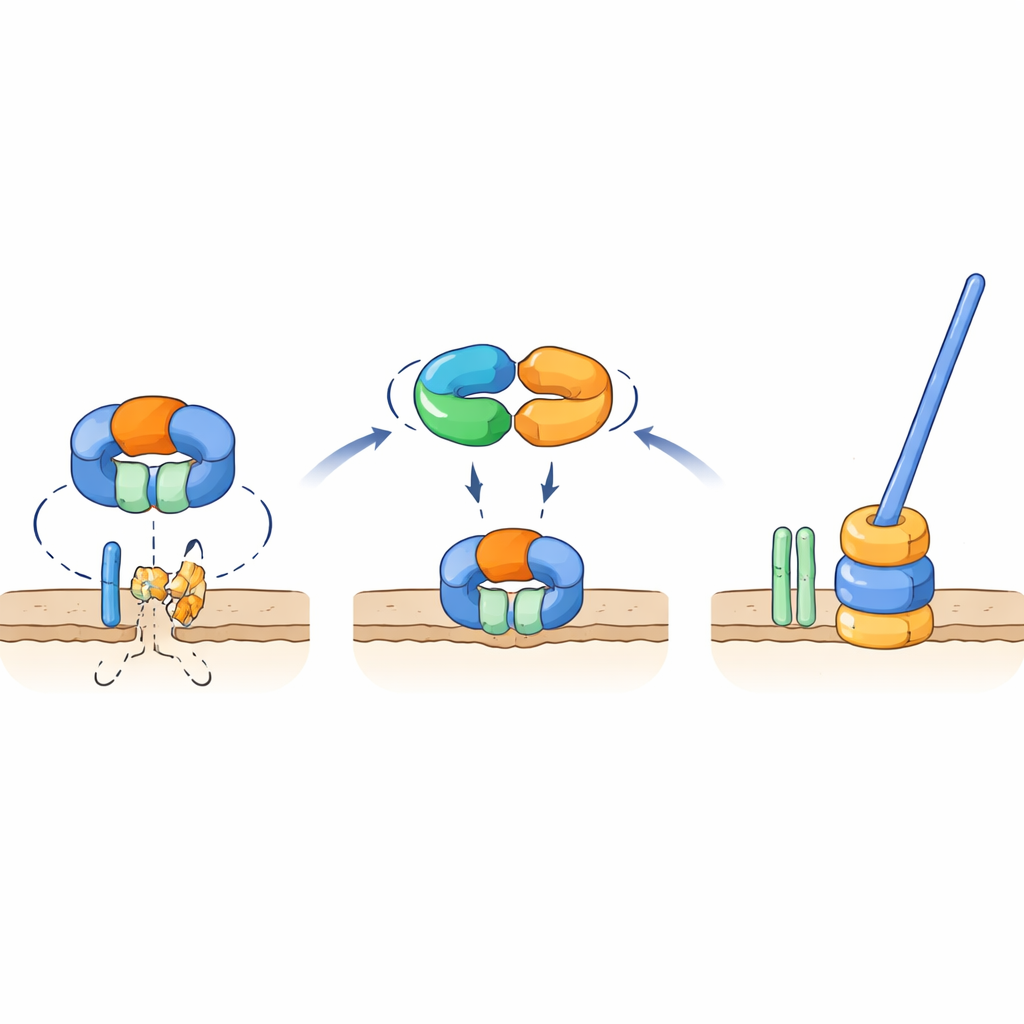
Een moleculaire schakel die een sleutelonderdeel van de flagel beschermt
Genetische en interactietesten toonden aan dat TrxA en PtxA fysiek aan elkaar binden, en dat het EKVEQ-motief essentieel is voor dit partnerschap. Met een bacterieel two-hybrid systeem vonden de onderzoekers dat TrxA en PtxA ook interageren met ClpC, een chaperonnecomponent van het Clp-proteasecomplex — een moleculaire machine die eiwitten ontvouwt en in een tonvormige “vernietiger” voert. Om te onderzoeken hoe dit met flagellen samenhangt, stelden ze de niet-beweeglijke TrxA- en PtxA-mutanten bloot aan UV-licht en selecteerden zeldzame suppressorstammen waarvan de sporen de beweeglijkheid terugkregen. Veel van deze suppressoren droegen mutaties in ClpC of in FliR, een membraaneiwit dat deel uitmaakt van de flagellaire exportpoort aan de basis van de staart. Het terugintroduceren van diezelfde mutaties in stammen zonder TrxA of PtxA herstelde de beweeglijkheid, en het verwijderen van fliR in een anders normale achtergrond maakte de flagellen helemaal ongedaan. Deze bevindingen ondersteunen een model waarbij, onder niet-flagellerende omstandigheden, het ClpC-bevattende protease FliR afbreekt en zo de assemblage van de flagellarbasis voorkomt. Wanneer de omstandigheden gunstig zijn voor flagelvorming, bindt het TrxA–PtxA-complex aan ClpC en dempt zo zijn proteolytische activiteit, zodat FliR kan ophopen en de exportpoort — en vervolgens de volledige flagel — kan worden opgebouwd.
Waarom dit belangrijk is voor microbieel leven en eiwitevolutie
Dit werk onthult een fijn afgestemd systeem waarmee een bacterie snel kan schakelen tussen een inactief spoor en een actieve zwemmer door één kwetsbaar onderdeel van de flagelmachine te bewaken. Het toont ook een thioredoxine die een taak uitvoert die niet afhangt van zijn gebruikelijke chemie: in plaats van als redoxkatalysator te fungeren, gebruikt TrxA een kort geconserveerd motief om een regulerend complex met PtxA te vormen en een protease te controleren. Zulke rolwisselingen benadrukken hoe bestaande eiwitfamilies tijdens de evolutie kunnen worden hergebruikt als nieuwe regulatiemodules, waardoor bacteriën zoals A. missouriensis complexe levenscyclusovergangen kunnen coördineren met slechts een handvol zorgvuldig geplaatste moleculaire interacties.
Bronvermelding: Kimura, T., Maeda, S., Suzuki, R. et al. Thioredoxin and its partner protein are essential for zoospore flagellar formation in Actinoplanes missouriensis. Commun Biol 9, 532 (2026). https://doi.org/10.1038/s42003-026-09784-8
Trefwoorden: bacteriële motiliteit, flagella-assemblage, eiwitregulatie, thioredoxine, proteasecontrole